

C/EBPalpha regulates generation of C/EBPbeta isoforms through activation of specific proteolytic cleavage
- PMID: 10022857
- PMCID: PMC83963
- DOI: 10.1128/MCB.19.3.1695
C/EBPalpha regulates generation of C/EBPbeta isoforms through activation of specific proteolytic cleavage
Abstract
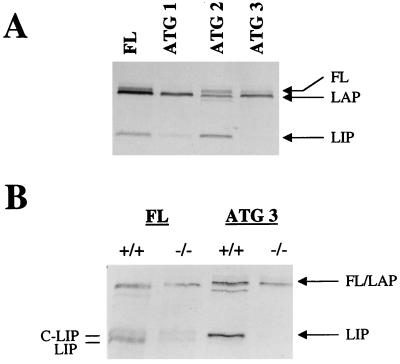
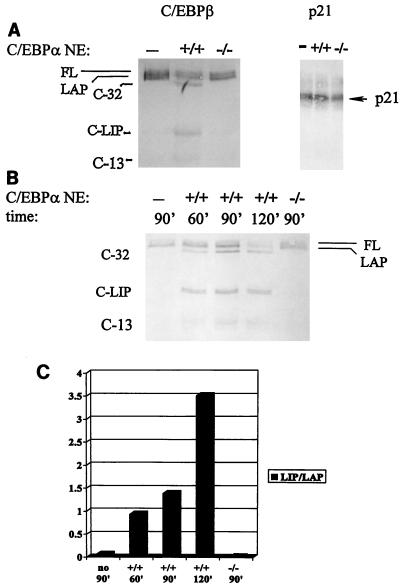
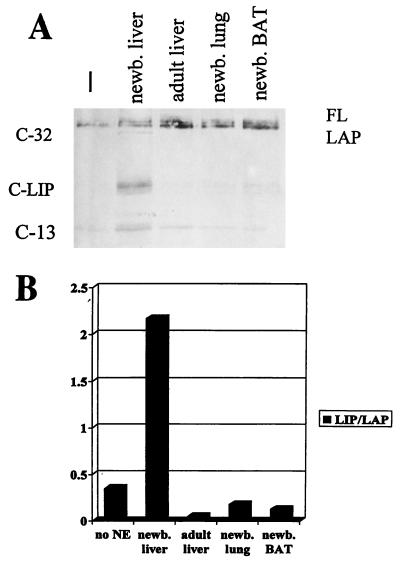
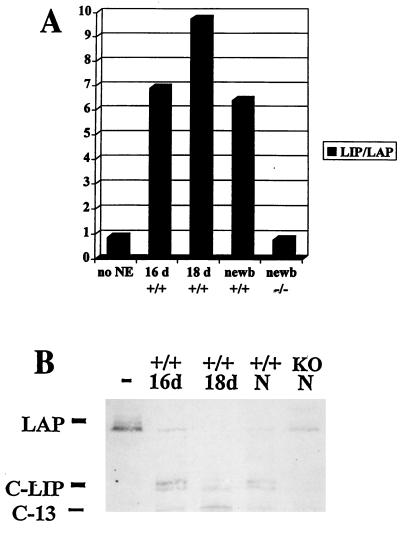
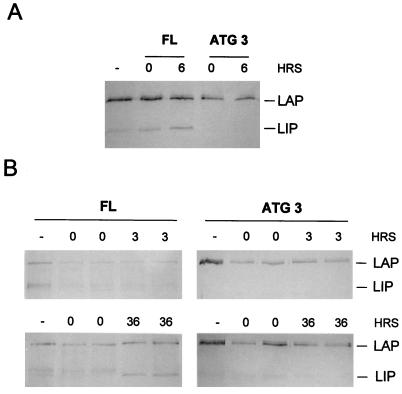
C/EBPalpha and C/EBPbeta are intronless genes that can produce several N-terminally truncated isoforms through the process of alternative translation initiation at downstream AUG codons. C/EBPbeta has been reported to produce four isoforms: full-length 38-kDa C/EBPbeta, 35-kDa LAP (liver-enriched transcriptional activator protein), 21-kDa LIP (liver-enriched transcriptional inhibitory protein), and a 14-kDa isoform. In this report, we investigated the mechanisms by which C/EBPbeta isoforms are generated in the liver and in cultured cells. Using an in vitro translation system, we found that LIP can be generated by two mechanisms: alternative translation and a novel mechanism-specific proteolytic cleavage of full-length C/EBPbeta. Studies of mice in which the C/EBPalpha gene had been deleted (C/EBPalpha-/-) showed that the regulation of C/EBPbeta proteolysis is dependent on C/EBPalpha. The induction of C/EBPalpha in cultured cells leads to induced cleavage of C/EBPbeta to generate the LIP isoform. We characterized the cleavage activity in mouse liver extracts and found that the proteolytic cleavage activity is specific to prenatal and newborn livers, is sensitive to chymostatin, and is completely abolished in C/EBPalpha-/- animals. The lack of cleavage activity in the livers of C/EBPalpha-/- mice correlates with the decreased levels of LIP in the livers of these animals. Analysis of LIP production during liver regeneration showed that, in this system, the transient induction of LIP is dependent on the third AUG codon and most likely involves translational control. We propose that there are two mechanisms by which C/EBPbeta isoforms might be generated in the liver and in cultured cells: one that is determined by translation and a second that involves C/EBPalpha-dependent, specific proteolytic cleavage of full-length C/EBPbeta. The latter mechanism implicates C/EBPalpha in the regulation of posttranslational generation of the dominant negative C/EBPbeta isoform, LIP.
Figures
Similar articles
-
CCAAT/enhancer binding protein alpha (C/EBPalpha) is an important mediator of mouse C/EBPbeta protein isoform production.Hepatology. 1999 Feb;29(2):597-601. doi: 10.1002/hep.510290245. Hepatology. 1999. PMID: 9918941
-
Translational induction of liver-enriched transcriptional inhibitory protein during acute phase response leads to repression of CCAAT/enhancer binding protein alpha mRNA.J Biol Chem. 2000 Sep 1;275(35):27406-13. doi: 10.1074/jbc.M002343200. J Biol Chem. 2000. PMID: 10854425
-
CUG repeat binding protein (CUGBP1) interacts with the 5' region of C/EBPbeta mRNA and regulates translation of C/EBPbeta isoforms.Nucleic Acids Res. 1999 Nov 15;27(22):4517-25. doi: 10.1093/nar/27.22.4517. Nucleic Acids Res. 1999. PMID: 10536163 Free PMC article.
-
CCAAT/enhancer binding proteins in normal mammary development and breast cancer.Breast Cancer Res. 2002;4(3):113-21. doi: 10.1186/bcr428. Epub 2002 Apr 19. Breast Cancer Res. 2002. PMID: 12052253 Free PMC article. Review.
-
CCAAT/enhancer-binding protein beta: its role in breast cancer and associations with receptor tyrosine kinases.Expert Rev Mol Med. 2009 Apr 8;11:e12. doi: 10.1017/S1462399409001033. Expert Rev Mol Med. 2009. PMID: 19351437 Free PMC article. Review.
Cited by
-
TSC1 controls IL-1β expression in macrophages via mTORC1-dependent C/EBPβ pathway.Cell Mol Immunol. 2016 Sep;13(5):640-50. doi: 10.1038/cmi.2015.43. Epub 2015 May 25. Cell Mol Immunol. 2016. PMID: 27593484 Free PMC article.
-
Regulation of CCAAT/enhancer-binding protein-beta isoform synthesis by alternative translational initiation at multiple AUG start sites.Nucleic Acids Res. 2001 Jul 15;29(14):3087-98. doi: 10.1093/nar/29.14.3087. Nucleic Acids Res. 2001. PMID: 11452034 Free PMC article.
-
An enhancer element 6 kb upstream of the mouse HNF4alpha1 promoter is activated by glucocorticoids and liver-enriched transcription factors.Nucleic Acids Res. 2001 Sep 1;29(17):3495-505. doi: 10.1093/nar/29.17.3495. Nucleic Acids Res. 2001. PMID: 11522818 Free PMC article.
-
S phase-specific proteolytic cleavage is required to activate stable DNA binding by the CDP/Cut homeodomain protein.Mol Cell Biol. 2001 Sep;21(18):6332-45. doi: 10.1128/MCB.21.18.6332-6345.2001. Mol Cell Biol. 2001. PMID: 11509674 Free PMC article.
-
The role of C/EBPbeta in mammary gland development and breast cancer.J Mammary Gland Biol Neoplasia. 2003 Apr;8(2):191-204. doi: 10.1023/a:1025900908026. J Mammary Gland Biol Neoplasia. 2003. PMID: 14635794 Review.
References
-
- Birkenmeier E H, Gwynn B, Howard S, Jerry J, Gordon J I, Landschulz W H, McKnight S L. Tissue-specific expression, developmental regulation, and genetic mapping of the gene encoding CCAAT/enhancer binding protein. Genes Dev. 1989;3:1146–1156. - PubMed
-
- Burgess-Beusse B L, Timchenko N A, Darlington G J. CCAAT/enhancer binding protein α (C/EBPα) is an important mediator of mouse C/EBPβ protein isoform production. Hepatology. 1999;29:597–601. - PubMed
-
- Cao Z, Umek R M, McKnight S L. Regulated expression of three C/EBP isoforms during adipose conversion of 3T3-L1 cells. Genes Dev. 1991;5:1538–1552. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials