

Substrate specificities of SR proteins in constitutive splicing are determined by their RNA recognition motifs and composite pre-mRNA exonic elements
- PMID: 10022872
- PMCID: PMC83978
- DOI: 10.1128/MCB.19.3.1853
Substrate specificities of SR proteins in constitutive splicing are determined by their RNA recognition motifs and composite pre-mRNA exonic elements
Abstract
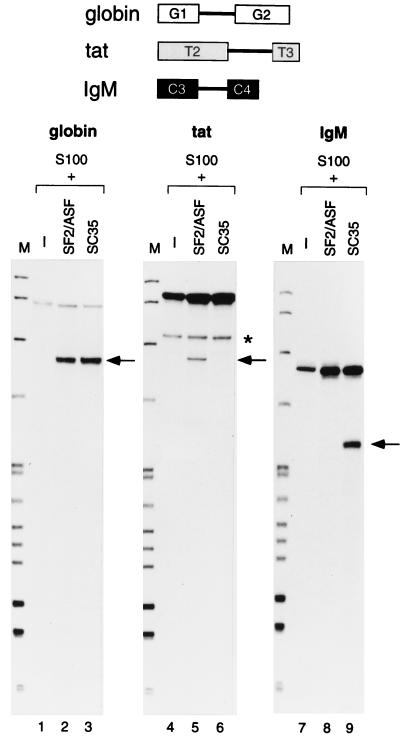
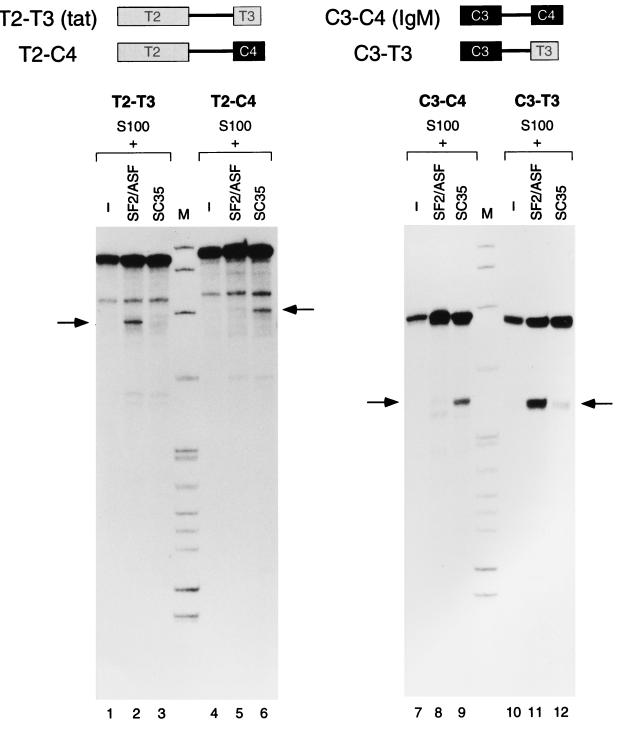
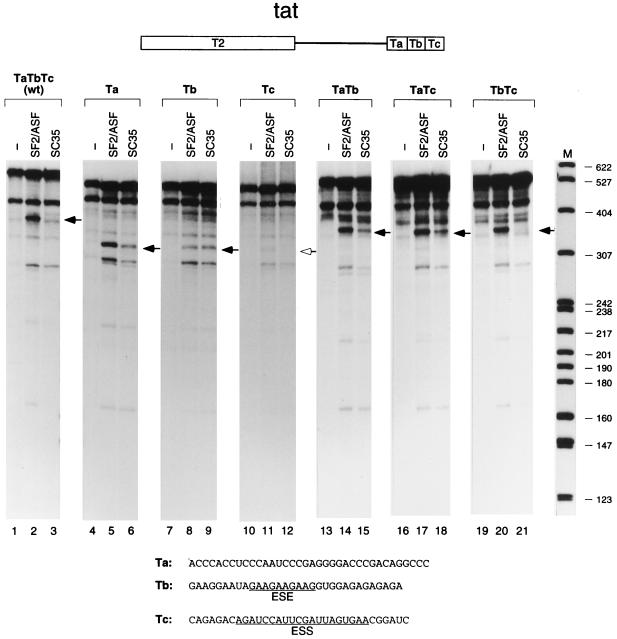
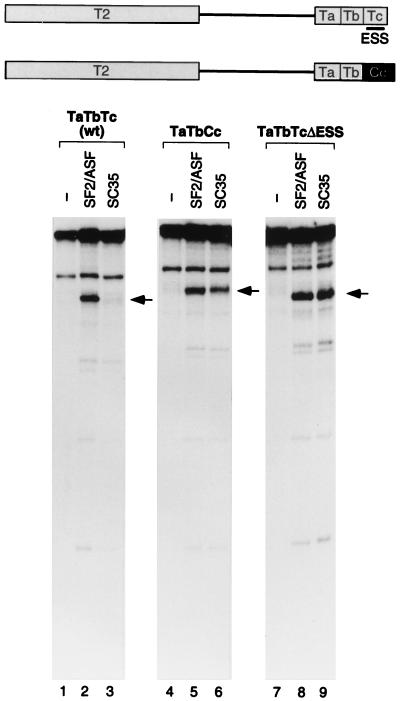
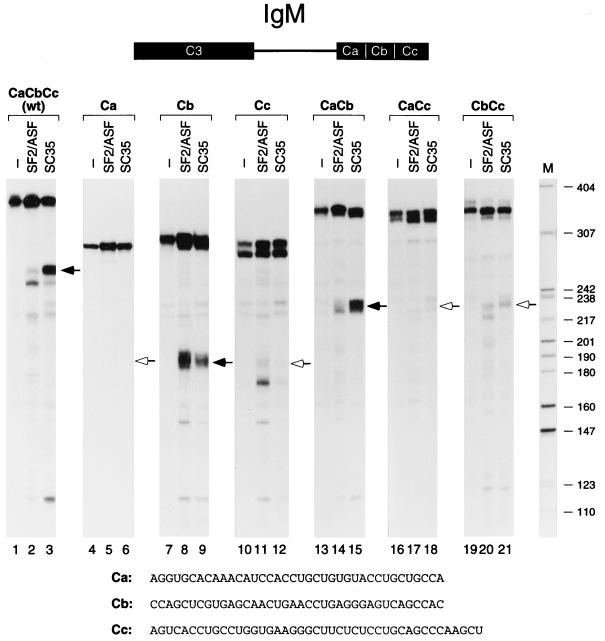
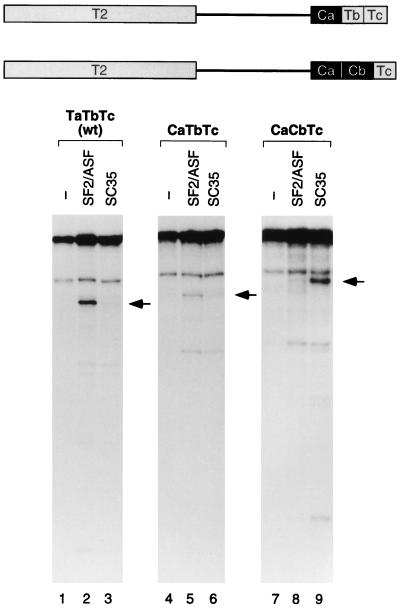
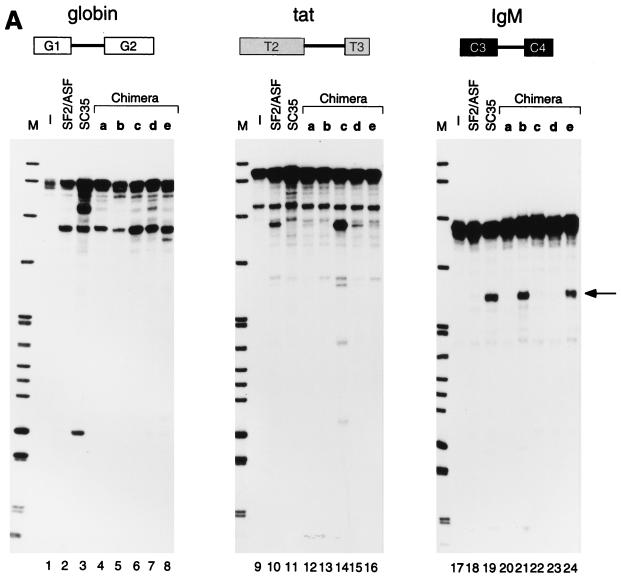
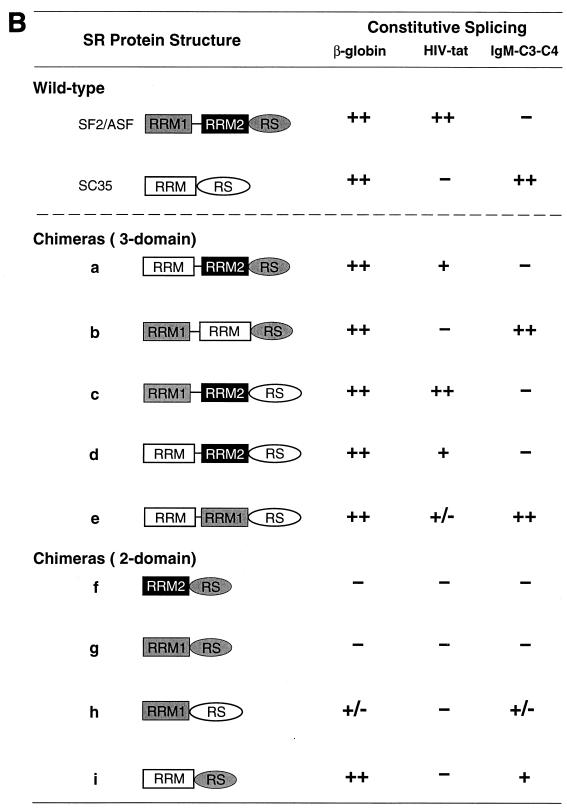
We report striking differences in the substrate specificities of two human SR proteins, SF2/ASF and SC35, in constitutive splicing. beta-Globin pre-mRNA (exons 1 and 2) is spliced indiscriminately with either SR protein. Human immunodeficiency virus tat pre-mRNA (exons 2 and 3) and immunoglobulin mu-chain (IgM) pre-mRNA (exons C3 and C4) are preferentially spliced with SF2/ASF and SC35, respectively. Using in vitro splicing with mutated or chimeric derivatives of the tat and IgM pre-mRNAs, we defined specific combinations of segments in the downstream exons, which mediate either positive or negative effects to confer SR protein specificity. A series of recombinant chimeric proteins consisting of domains of SF2/ASF and SC35 in various combinations was used to localize trans-acting domains responsible for substrate specificity. The RS domains of SF2/ASF and SC35 can be exchanged without effect on substrate specificity. The RNA recognition motifs (RRMs) of SF2/ASF are active only in the context of a two-RRM structure, and RRM2 has a dominant role in substrate specificity. In contrast, the single RRM of SC35 can function alone, but its substrate specificity can be influenced by the presence of an additional RRM. The RRMs behave as modules that, when present in different combinations, can have positive, neutral, or negative effects on splicing, depending upon the specific substrate. We conclude that SR protein-specific recognition of specific positive and negative pre-mRNA exonic elements via one or more RRMs is a crucial determinant of the substrate specificity of SR proteins in constitutive splicing.
Figures
References
-
- Cáceres J F, Krainer A R. Mammalian pre-mRNA splicing factors. In: Krainer A R, editor. Eukaryotic mRNA processing. Oxford, United Kingdom: Oxford University Press; 1997. pp. 174–212.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous