

Phosphorylation of the cap-binding protein eukaryotic translation initiation factor 4E by protein kinase Mnk1 in vivo
- PMID: 10022874
- PMCID: PMC83980
- DOI: 10.1128/MCB.19.3.1871
Phosphorylation of the cap-binding protein eukaryotic translation initiation factor 4E by protein kinase Mnk1 in vivo
Abstract
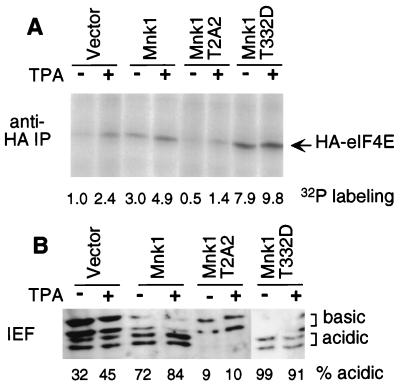
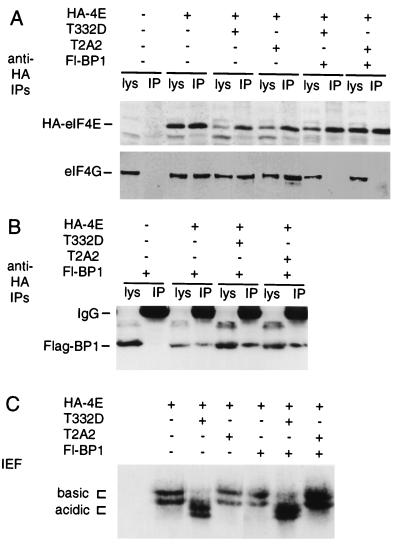
Eukaryotic translation initiation factor 4E (eIF4E) binds to the mRNA 5' cap and brings the mRNA into a complex with other protein synthesis initiation factors and ribosomes. The activity of mammalian eIF4E is important for the translation of capped mRNAs and is thought to be regulated by two mechanisms. First, eIF4E is sequestered by binding proteins, such as 4EBP1, in quiescent cells. Mitogens induce the release of eIF4E by stimulating the phosphorylation of 4EBP1. Second, mitogens and stresses induce the phosphorylation of eIF4E at Ser 209, increasing the affinity of eIF4E for capped mRNA and for an associated scaffolding protein, eIF4G. We previously showed that a mitogen- and stress-activated kinase, Mnk1, phosphorylates eIF4E in vitro at the physiological site. Here we show that Mnk1 regulates eIF4E phosphorylation in vivo. Mnk1 binds directly to eIF4G and copurifies with eIF4G and eIF4E. We identified activating phosphorylation sites in Mnk1 and developed dominant-negative and activated mutants. Expression of dominant-negative Mnk1 reduces mitogen-induced eIF4E phosphorylation, while expression of activated Mnk1 increases basal eIF4E phosphorylation. Activated mutant Mnk1 also induces extensive phosphorylation of eIF4E in cells overexpressing 4EBP1. This suggests that phosphorylation of eIF4E is catalyzed by Mnk1 or a very similar kinase in cells and is independent of other mitogenic signals that release eIF4E from 4EBP1.
Figures
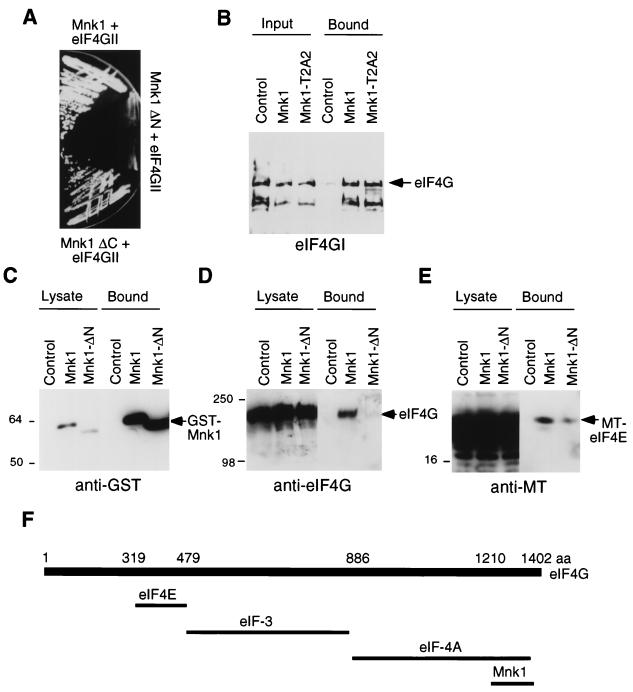
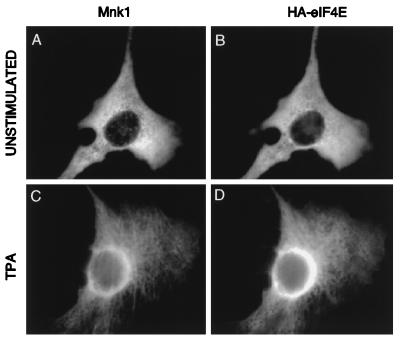
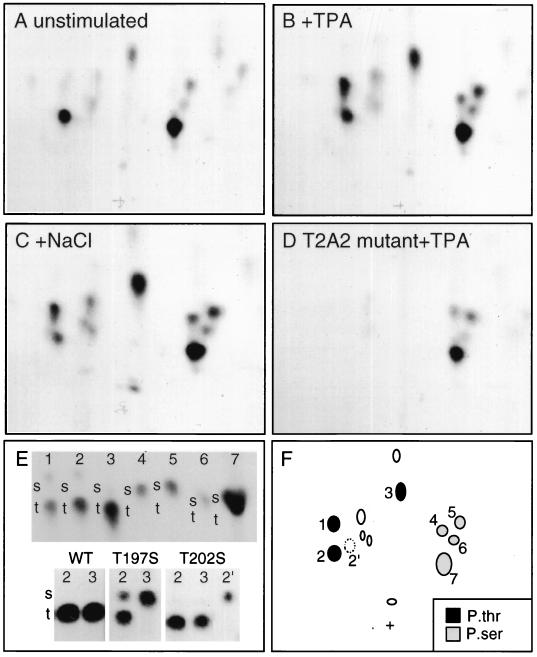
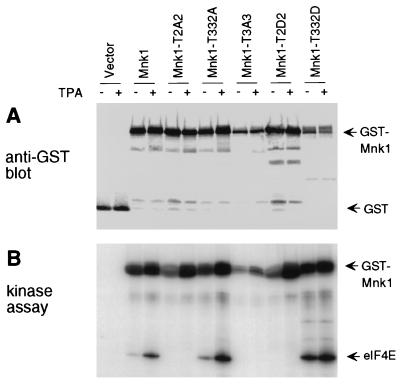
References
-
- Alvarez E, Northwood I C, Gonzalez E A, Latour D A, Seth A, Abate C, Curran T. Pro-Leu-Ser/Thr-Pro is a consensus primary sequence for substrate phosphorylation. Characterization of the phosphorylation of c-myc and c-jun proteins by an epidermal growth factor receptor threonine 669 kinase. J Biol Chem. 1991;266:15277–15285. - PubMed
-
- Boyle W J, van der Geer P, Hunter T. Phosphopeptide mapping and phosphoamino acid analysis by two-dimensional separation on thin-layer cellulose plates. Methods Enzymol. 1991;201:110–148. - PubMed
-
- Brunn G J, Hudson C C, Sekulic A, Williams J M, Hosoi H, Houghton P J, Lawrence J C, Jr, Abraham R T. Phosphorylation of the translational repressor PHAS-I by the mammalian target of rapamycin. Science. 1997;277:99–101. - PubMed
-
- Bu X, Haas D W, Hagedorn C H. Novel phosphorylation sites of eukaryotic initiation factor-4F and evidence that phosphorylation stabilizes interactions of the p25 and p220 subunits. J Biol Chem. 1993;268:4975–4978. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous