

Reactive oxygen intermediate-dependent NF-kappaB activation by interleukin-1beta requires 5-lipoxygenase or NADPH oxidase activity
- PMID: 10022882
- PMCID: PMC83988
- DOI: 10.1128/MCB.19.3.1950
Reactive oxygen intermediate-dependent NF-kappaB activation by interleukin-1beta requires 5-lipoxygenase or NADPH oxidase activity
Abstract
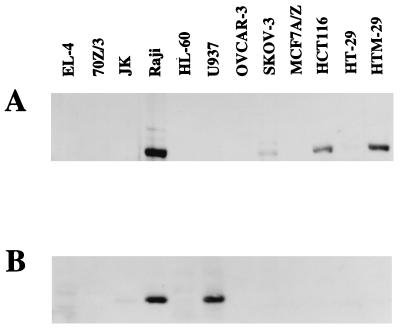
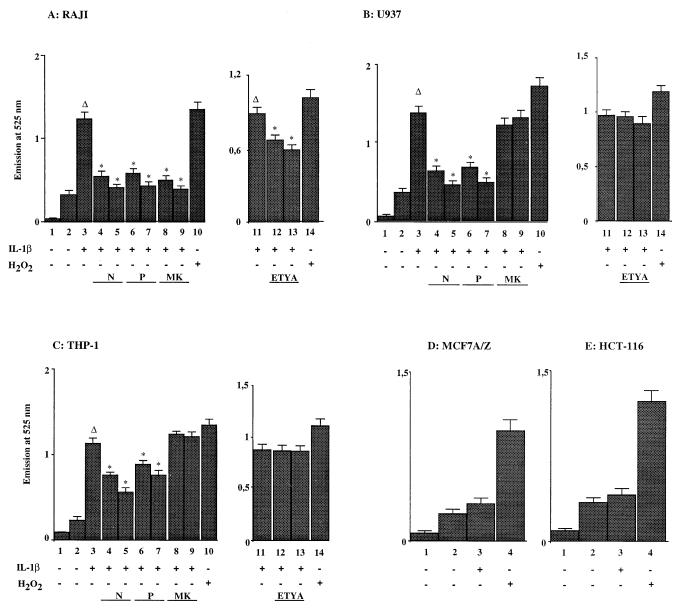
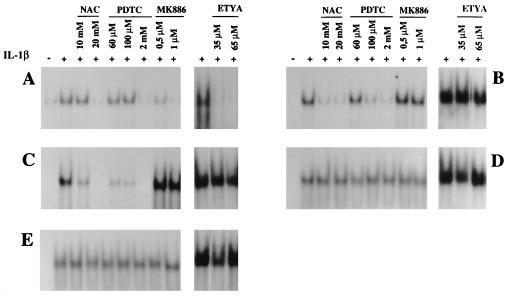
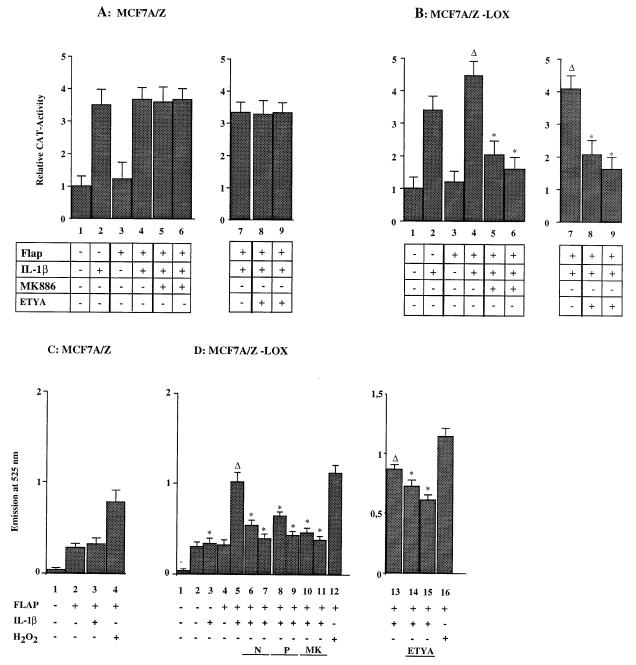
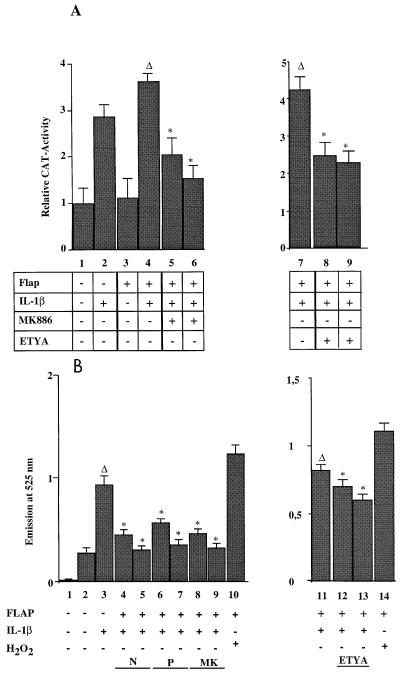
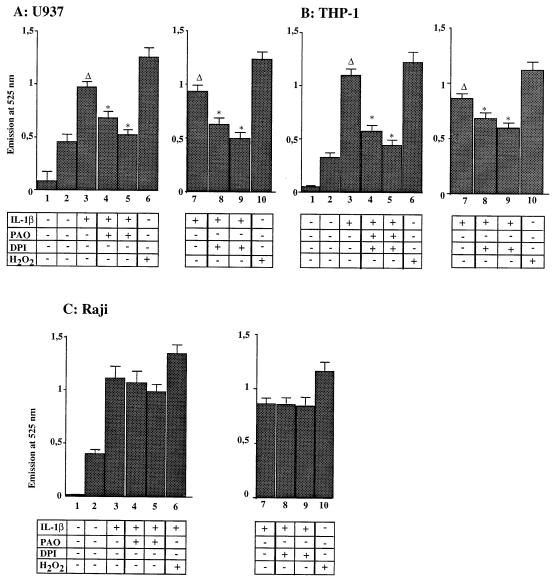
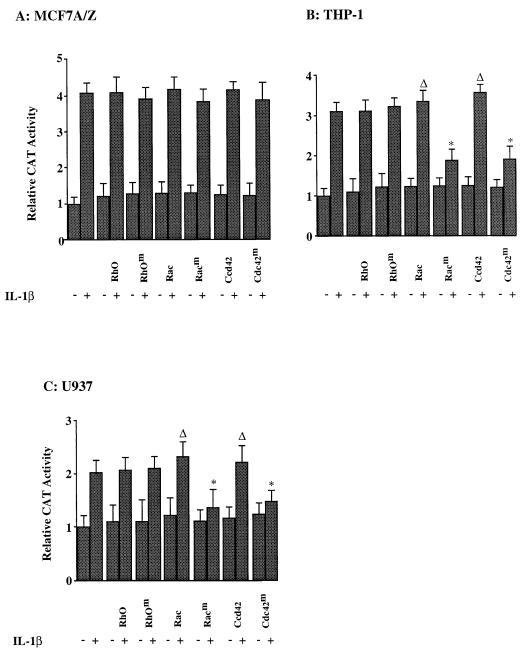
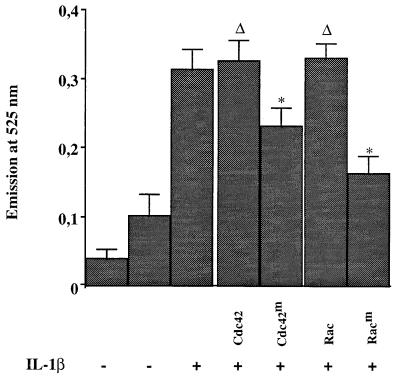
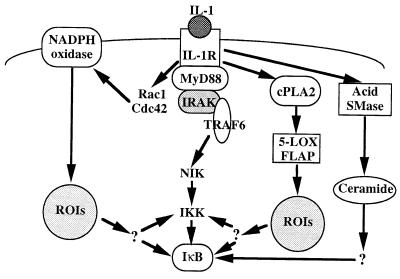
We previously reported that the role of reactive oxygen intermediates (ROIs) in NF-kappaB activation by proinflammatory cytokines was cell specific. However, the sources for ROIs in various cell types are yet to be determined and might include 5-lipoxygenase (5-LOX) and NADPH oxidase. 5-LOX and 5-LOX activating protein (FLAP) are coexpressed in lymphoid cells but not in monocytic or epithelial cells. Stimulation of lymphoid cells with interleukin-1beta (IL-1beta) led to ROI production and NF-kappaB activation, which could both be blocked by antioxidants or FLAP inhibitors, confirming that 5-LOX was the source of ROIs and was required for NF-kappaB activation in these cells. IL-1beta stimulation of epithelial cells did not generate any ROIs and NF-kappaB induction was not influenced by 5-LOX inhibitors. However, reintroduction of a functional 5-LOX system in these cells allowed ROI production and 5-LOX-dependent NF-kappaB activation. In monocytic cells, IL-1beta treatment led to a production of ROIs which is independent of the 5-LOX enzyme but requires the NADPH oxidase activity. This pathway involves the Rac1 and Cdc42 GTPases, two enzymes which are not required for NF-kappaB activation by IL-1beta in epithelial cells. In conclusion, three different cell-specific pathways lead to NF-kappaB activation by IL-1beta: a pathway dependent on ROI production by 5-LOX in lymphoid cells, an ROI- and 5-LOX-independent pathway in epithelial cells, and a pathway requiring ROI production by NADPH oxidase in monocytic cells.
Figures

References
-
- Abo A, Boyhan A, West I, Thrasher A J, Segal A W. Reconstitution of neutrophil NADPH oxidase activity in the cell-free system by four components: p67-phox, p47-phox, p21rac1 and cytochrome b-245. J Biol Chem. 1992;267:16767–16770. - PubMed
-
- Baldwin A S. The NF-κB and IκB proteins: new discoveries and insights. Annu Rev Immunol. 1996;14:649–681. - PubMed
-
- Barnes P J, Karin M. Nuclear factor-κB—a pivotal transcription factor in chronic inflammatory diseases. N Engl J Med. 1997;336:1066–1071. - PubMed
-
- Bokoch G M. Regulation of the phagocyte respiratory burst by small GTP-binding proteins. Trends Cell Biol. 1995;5:109–113. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous