

Progesterone inhibits estrogen-induced cyclin D1 and cdk4 nuclear translocation, cyclin E- and cyclin A-cdk2 kinase activation, and cell proliferation in uterine epithelial cells in mice
- PMID: 10022912
- PMCID: PMC84018
- DOI: 10.1128/MCB.19.3.2251
Progesterone inhibits estrogen-induced cyclin D1 and cdk4 nuclear translocation, cyclin E- and cyclin A-cdk2 kinase activation, and cell proliferation in uterine epithelial cells in mice
Abstract
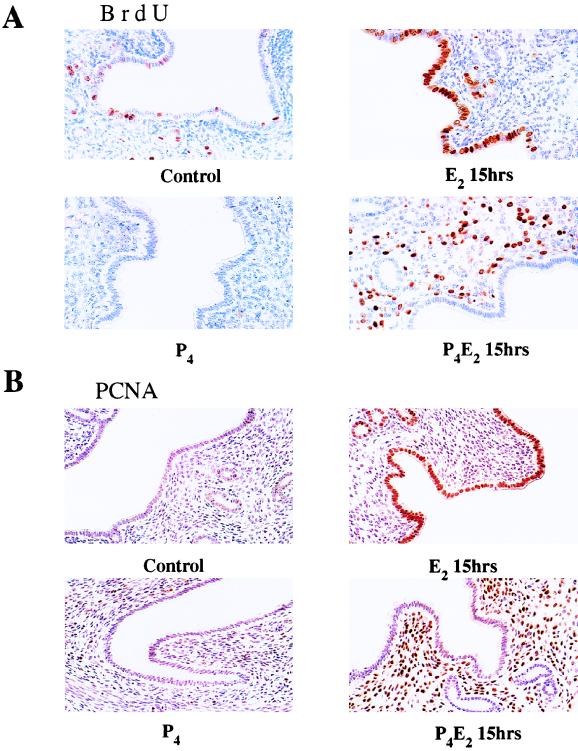
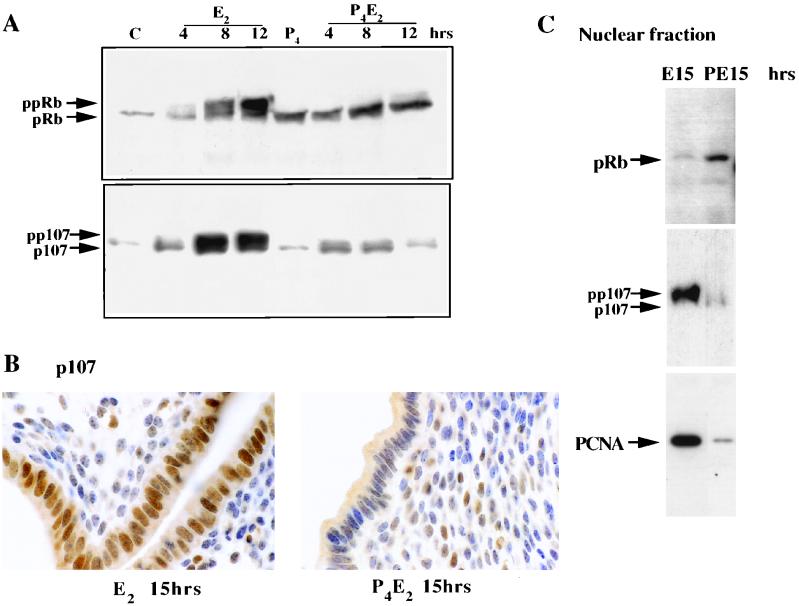
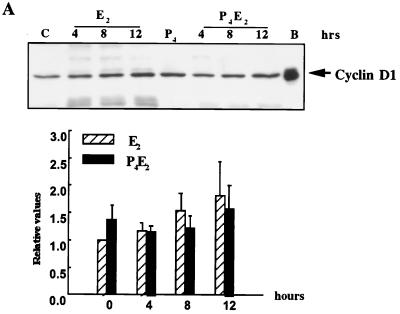
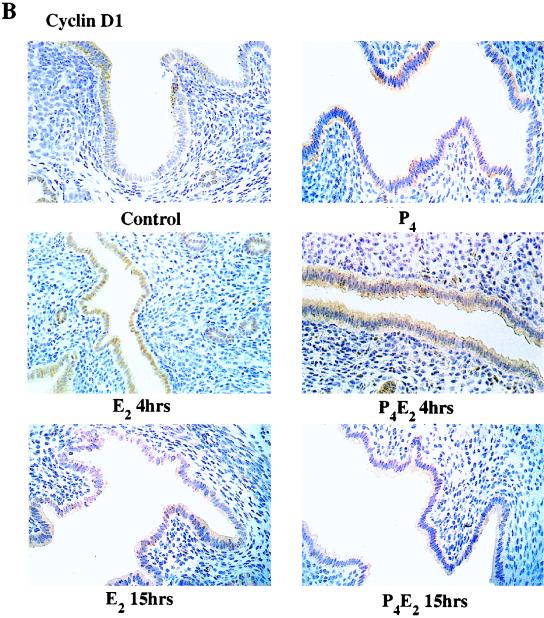
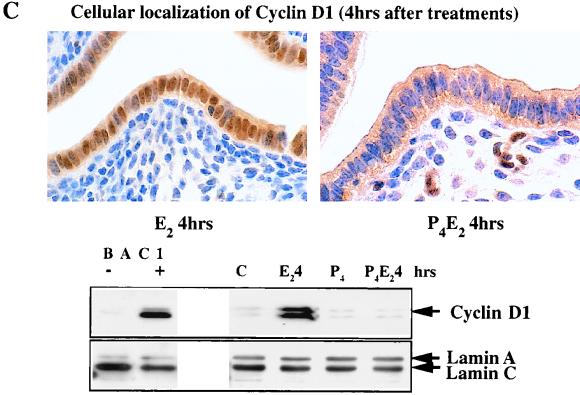
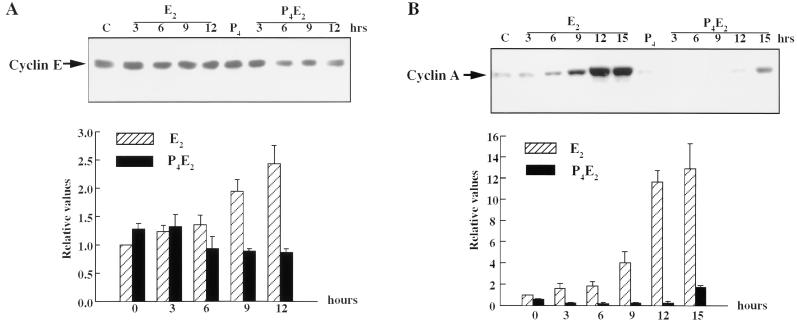
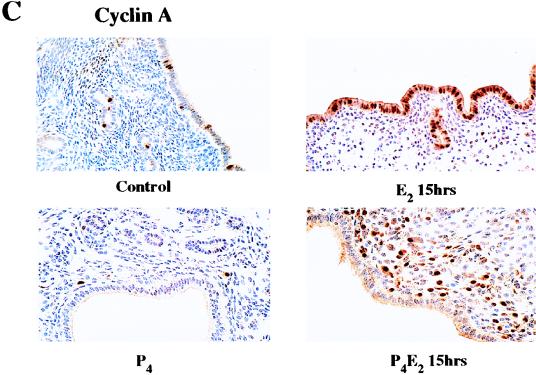
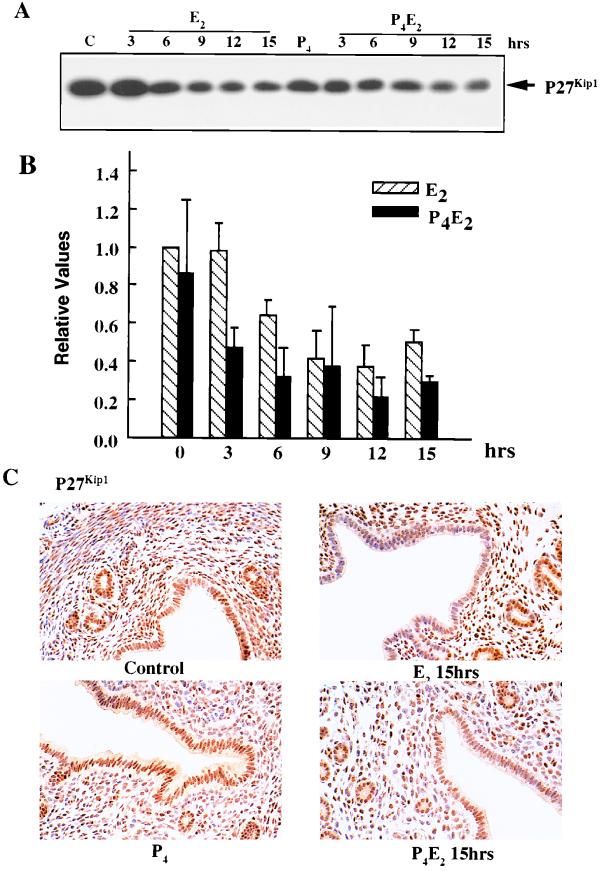
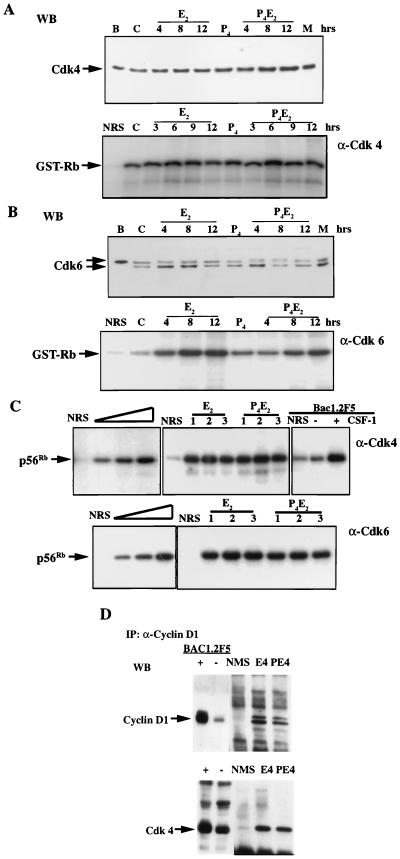
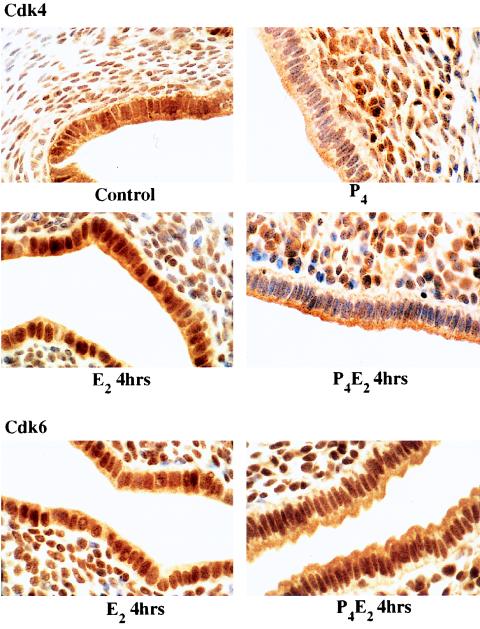
The response of the uterine epithelium to female sex steroid hormones provides an excellent model to study cell proliferation in vivo since both stimulation and inhibition of cell proliferation can be studied. Thus, when administered to ovariectomized adult mice 17beta-estradiol (E2) stimulates a synchronized wave of DNA synthesis and cell division in the epithelial cells, while pretreatment with progesterone (P4) completely inhibits this E2-induced cell proliferation. Using a simple method to isolate the uterine epithelium with high purity, we have shown that E2 treatment induces a relocalization of cyclin D1 and, to a lesser extent, cdk4 from the cytoplasm into the nucleus and results in the orderly activation of cyclin E- and cyclin A-cdk2 kinases and hyperphosphorylation of pRb and p107. P4 pretreatment did not alter overall levels of cyclin D1, cdk4, or cdk6 nor their associated kinase activities but instead inhibited the E2-induced nuclear localization of cyclin D1 to below the control level and, to a lesser extent, nuclear cdk4 levels, with a consequent inhibition of pRb and p107 phosphorylation. In addition, it abrogated E2-induced cyclin E-cdk2 activation by dephosphorylation of cdk2, followed by inhibition of cyclin A expression and consequently of cyclin A-cdk2 kinase activity and further inhibition of phosphorylation of pRb and p107. P4 is used therapeutically to oppose the effect of E2 during hormone replacement therapy and in the treatment of uterine adenocarcinoma. This study showing a novel mechanism of cell cycle inhibition by P4 may provide the basis for the development of new antiestrogens.
Figures

Similar articles
-
Disruption of the actin cytoskeleton leads to inhibition of mitogen-induced cyclin E expression, Cdk2 phosphorylation, and nuclear accumulation of the retinoblastoma protein-related p107 protein.Exp Cell Res. 2000 Aug 25;259(1):35-53. doi: 10.1006/excr.2000.4966. Exp Cell Res. 2000. PMID: 10942577
-
Cyclin D2 compensates for the loss of cyclin D1 in estrogen-induced mouse uterine epithelial cell proliferation.Mol Endocrinol. 2003 Jul;17(7):1368-81. doi: 10.1210/me.2003-0036. Epub 2003 Mar 20. Mol Endocrinol. 2003. PMID: 12649329
-
Mechanisms of cyclin-dependent kinase inactivation by progestins.Mol Cell Biol. 1998 Apr;18(4):1812-25. doi: 10.1128/MCB.18.4.1812. Mol Cell Biol. 1998. PMID: 9528753 Free PMC article.
-
Ras regulation of cyclin-dependent immunoprecipitation kinase assays.Methods Enzymol. 2001;333:127-38. doi: 10.1016/s0076-6879(01)33051-3. Methods Enzymol. 2001. PMID: 11400330 Review. No abstract available.
-
Mammalian cyclin-dependent kinases.Trends Biochem Sci. 2005 Nov;30(11):630-41. doi: 10.1016/j.tibs.2005.09.005. Epub 2005 Oct 19. Trends Biochem Sci. 2005. PMID: 16236519 Review.
Cited by
-
Estradiol-17beta regulates mouse uterine epithelial cell proliferation through insulin-like growth factor 1 signaling.Proc Natl Acad Sci U S A. 2007 Oct 2;104(40):15847-51. doi: 10.1073/pnas.0705749104. Epub 2007 Sep 25. Proc Natl Acad Sci U S A. 2007. PMID: 17895382 Free PMC article.
-
Estrogen stimulates expression of p21Waf1/Cip1 in mouse uterine luminal epithelium.Endocrine. 2002 Apr;17(3):233-9. doi: 10.1385/ENDO:17:3:233. Endocrine. 2002. PMID: 12108525
-
A mouse model to dissect progesterone signaling in the female reproductive tract and mammary gland.Genesis. 2010 Feb;48(2):106-13. doi: 10.1002/dvg.20586. Genesis. 2010. PMID: 20029965 Free PMC article.
-
Estradiol and progesterone exhibit similar patterns of hepatic gene expression regulation in the bovine model.PLoS One. 2013 Sep 17;8(9):e73552. doi: 10.1371/journal.pone.0073552. eCollection 2013. PLoS One. 2013. PMID: 24069207 Free PMC article.
-
Identification and characterization of progesterone- and estrogen-regulated MicroRNAs in mouse endometrial epithelial cells.Reprod Sci. 2015 Feb;22(2):223-34. doi: 10.1177/1933719114537714. Epub 2014 Jun 12. Reprod Sci. 2015. PMID: 24925854 Free PMC article.
References
-
- Altucci L, Addeo R, Cicatiello L, Dauvois S, Parker M G, Truss M, Beato M, Sica V, Bresciani F, Weisz A. 17β-Estradiol induces cyclin D1 gene transcription, p36D1-p34cdk4 complex activation and p105Rb phosphorylation during mitogenic stimulation of G1-arrested human breast cancer cells. Oncogene. 1996;12:2315–2324. - PubMed
-
- Altucci L, Addeo R, Cicatiello L, Germano D, Pacilio C, Battista T, Cancemi M, Petrizzi V B, Bresciani F, Weisz A. Estrogen induces early and timed activation of cyclin-dependent kinases 4, 5, and 6 and increases cyclin messenger ribonucleic acid expression in rat uterus. Endocrinology. 1997;138:978–984. - PubMed
-
- Baldin V, Lukas J, Marcote M, Pagano M, Draetta G. Cyclin D1 is a nuclear protein required for cell cycle progression in G1. Genes Dev. 1993;7:812–821. - PubMed
-
- Castles C G, Fuqua S A W. Alterations within the estrogen receptor in breast cancer. In: Pasqualini J R, Katzenellenbogen B S, editors. Hormonal-dependent cancer. New York, N.Y: Marcel Dekker, Inc.; 1996. pp. 81–105.
-
- Cheng M, Diehl J A, Fero M, Olivier P, Randel E, Roberts J M, Sherr C J, Roussel M F. Fourteenth Annual Meeting on Oncogenes. (Abstract.) 1998. Cdk inhibitors p27Kip1 and p21Cip1 are required for assembly of active cyclin D-dependent kinases in mouse fibroblasts; p. 39.
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials