

The yeast ser/thr phosphatases sit4 and ppz1 play opposite roles in regulation of the cell cycle
- PMID: 10022927
- PMCID: PMC84033
- DOI: 10.1128/MCB.19.3.2408
The yeast ser/thr phosphatases sit4 and ppz1 play opposite roles in regulation of the cell cycle
Abstract
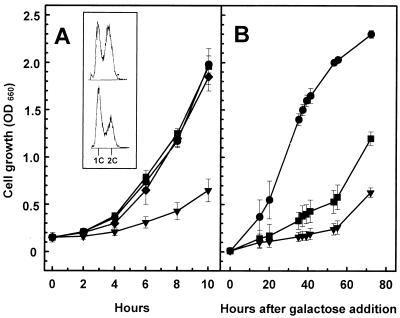
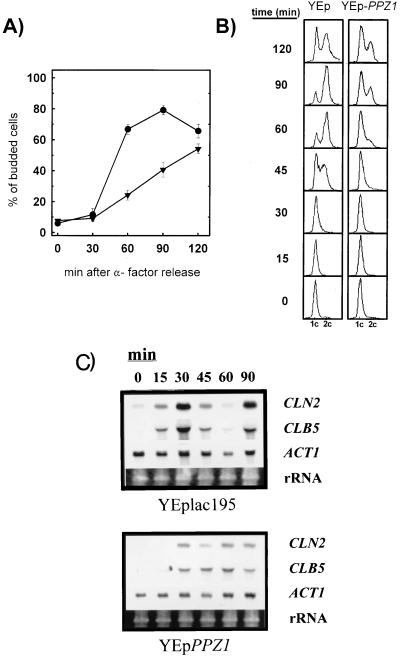
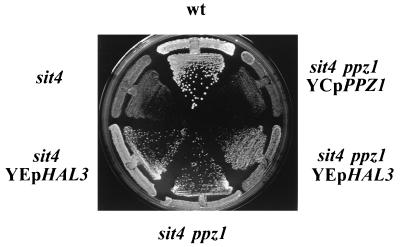
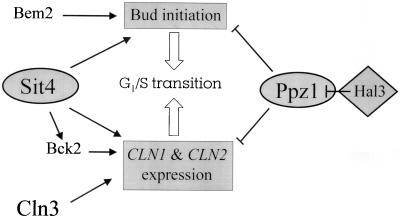
Yeast cells overexpressing the Ser/Thr protein phosphatase Ppz1 display a slow-growth phenotype. These cells recover slowly from alpha-factor or nutrient depletion-induced G1 arrest, showing a considerable delay in bud emergence as well as in the expression of the G1 cyclins Cln2 and Clb5. Therefore, an excess of the Ppz1 phosphatase interferes with the normal transition from G1 to S phase. The growth defect is rescued by overexpression of the HAL3/SIS2 gene, encoding a negative regulator of Ppz1. High-copy-number expression of HAL3/SIS2 has been reported to improve cell growth and to increase expression of G1 cyclins in sit4 phosphatase mutants. We show here that the described effects of HAL3/SIS2 on sit4 mutants are fully mediated by the Ppz1 phosphatase. The growth defect caused by overexpression of PPZ1 is intensified in strains with low G1 cyclin levels (such as bck2Delta or cln3Delta mutants), whereas mutation of PPZ1 rescues the synthetic lethal phenotype of sit4 cln3 mutants. These results reveal a role for Ppz1 as a regulatory component of the yeast cell cycle, reinforce the notion that Hal3/Sis2 serves as a negative modulator of the biological functions of Ppz1, and indicate that the Sit4 and Ppz1 Ser/Thr phosphatases play opposite roles in control of the G1/S transition.
Figures
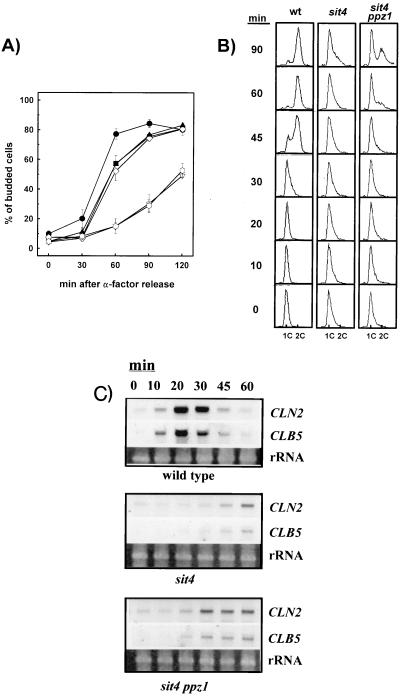
 ) were arrested in G1 phase by incubation with α-factor and released from arrest as indicated. Cells were taken at the indicated times, and the budding index was determined by microscopic counting. Data are means ± SEM of six experiments. (B) Analysis of DNA content of wild-type (wt), sit4 (strain JA-110), and sit4 ppz1 (strain A-112) cells by flow cytometry. Cells were arrested as described above, and samples were taken at different times after release from α-factor arrest. (C) The indicated strains were released form α-factor arrest, and samples were taken at various times. Total RNA was prepared, electrophoresed, transferred to membranes, and hybridized with the indicated probes. Ethidium bromide staining of rRNAs is shown for comparison.
) were arrested in G1 phase by incubation with α-factor and released from arrest as indicated. Cells were taken at the indicated times, and the budding index was determined by microscopic counting. Data are means ± SEM of six experiments. (B) Analysis of DNA content of wild-type (wt), sit4 (strain JA-110), and sit4 ppz1 (strain A-112) cells by flow cytometry. Cells were arrested as described above, and samples were taken at different times after release from α-factor arrest. (C) The indicated strains were released form α-factor arrest, and samples were taken at various times. Total RNA was prepared, electrophoresed, transferred to membranes, and hybridized with the indicated probes. Ethidium bromide staining of rRNAs is shown for comparison.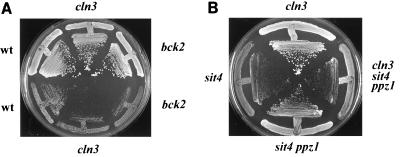

References
-
- Arndt K T, Styles C A, Fink G R. A suppressor of a HIS4 transcriptional defect encodes a protein with homology to the catalytic subunit of protein phosphatases. Cell. 1989;56:527–537. - PubMed
-
- Balcells L, Gómez N, Casamayor A, Clotet J, Ariño J. Regulation of salt tolerance in fission yeast by a protein-phosphatase-Z-like Ser/Thr protein phosphatase. Eur J Biochem. 1997;250:2476–2483. - PubMed
-
- Bastians H, Ponstingl H. The novel human protein serine/threonine phosphatase 6 is a functional homologue of budding yeast Sit4p and fission yeast ppe1, which are involved in cell cycle regulation. J Cell Sci. 1996;109:2865–2874. - PubMed
-
- Berben G, Dumont J, Gilliquet V, Bolle P, Hilger F. The YDp plasmids: a uniform set of vectors bearing versatile gene disruption cassettes for Saccharomyces cerevisiae. Yeast. 1991;7:475–477. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases