

Developmental synaptic changes increase the range of integrative capabilities of an identified excitatory neocortical connection
- PMID: 10024344
- PMCID: PMC6782183
- DOI: 10.1523/JNEUROSCI.19-05-01566.1999
Developmental synaptic changes increase the range of integrative capabilities of an identified excitatory neocortical connection
Abstract
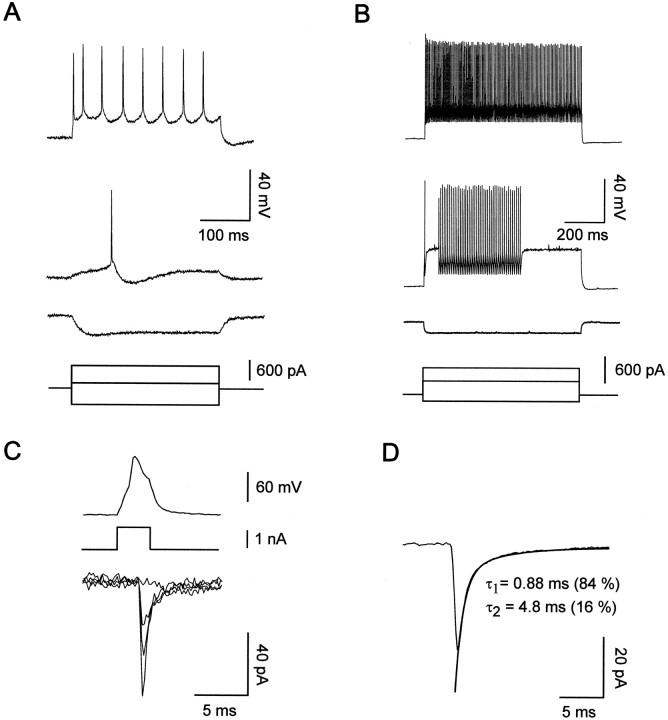
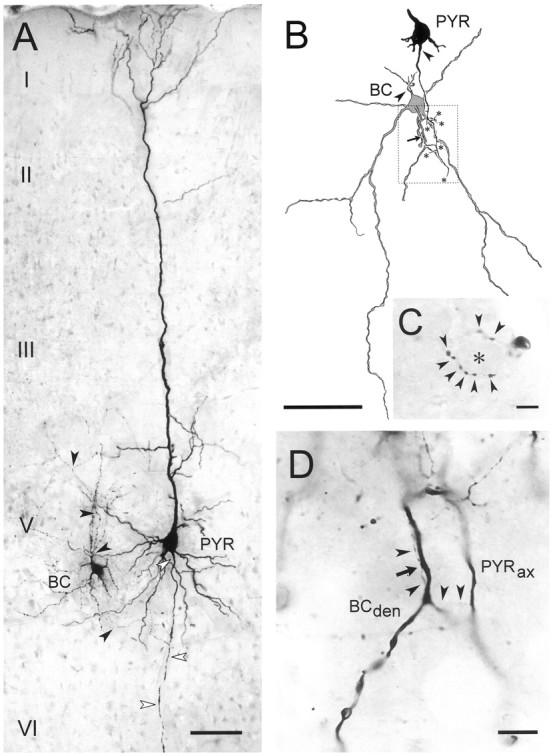
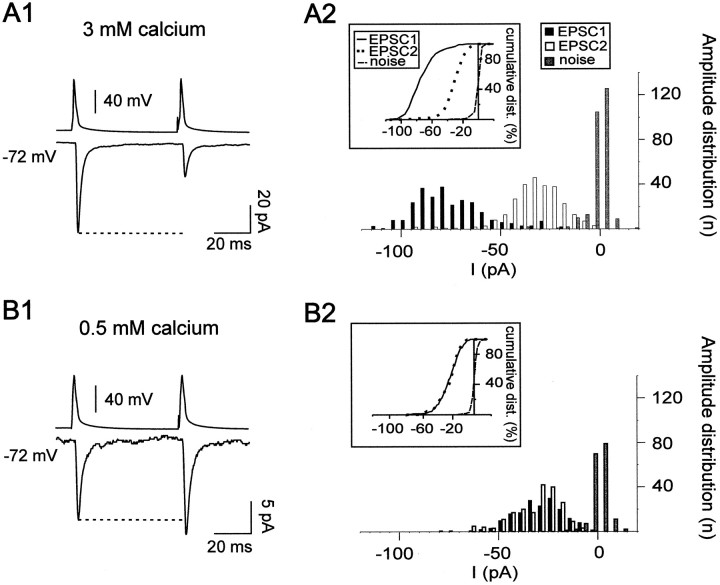
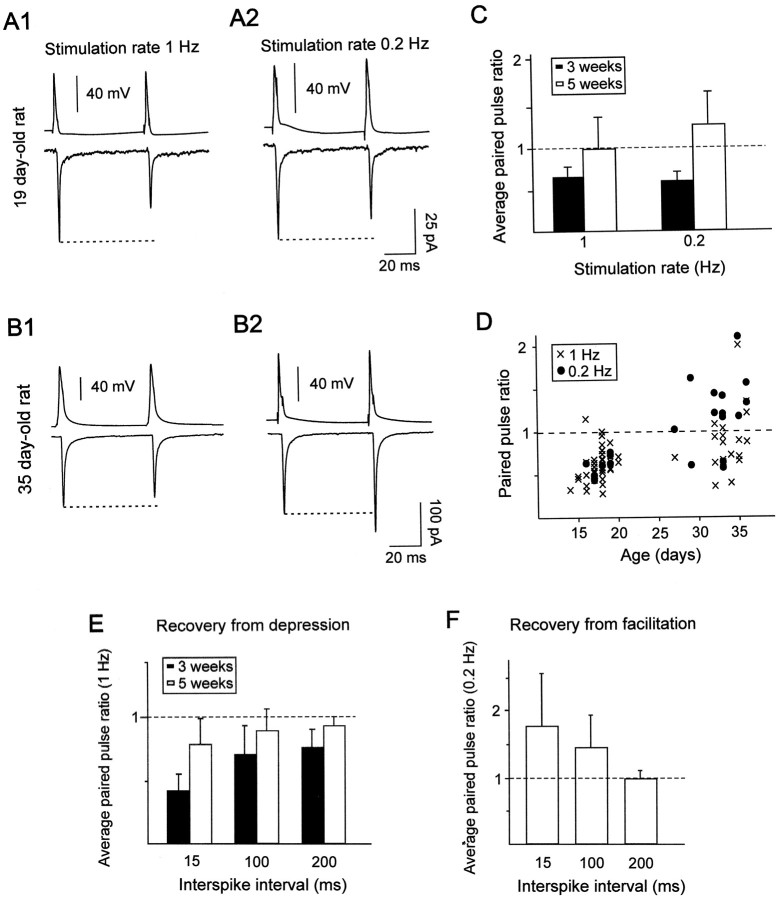
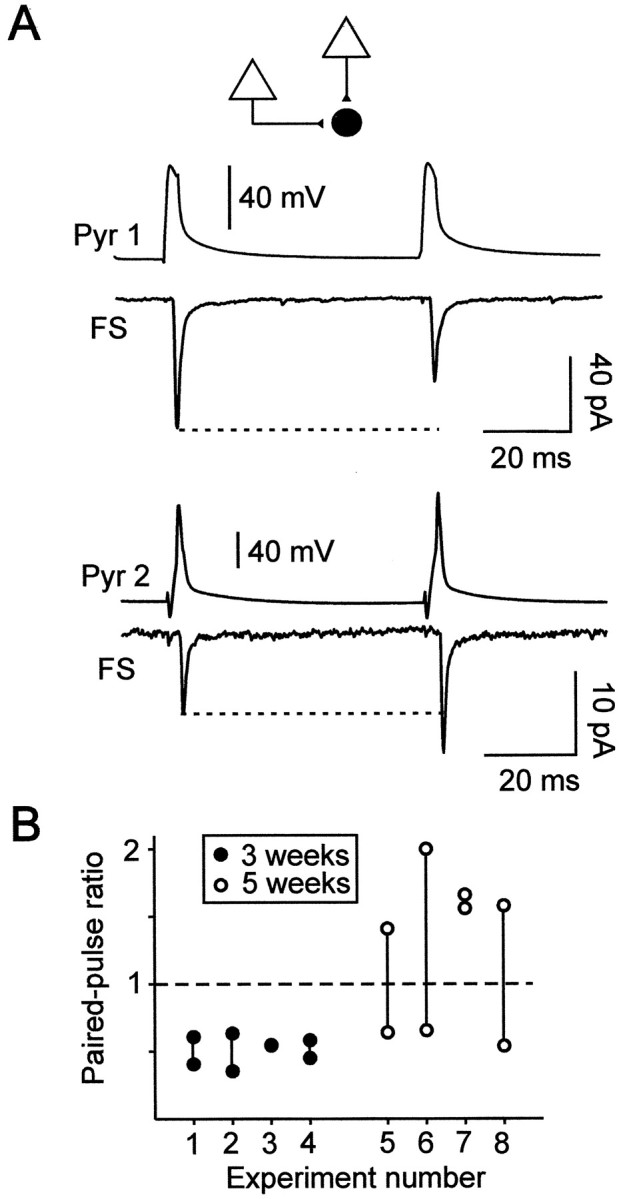
Excitatory synaptic transmission between pyramidal cells and fast-spiking (FS) interneurons of layer V of the motor cortex was investigated in acute slices by using paired recordings at 30 degrees C combined with morphological analysis. The presynaptic and postsynaptic properties at these identified central synapses were compared between 3- and 5-week-old rats. At these two postnatal developmental stages, unitary EPSCs were mediated by the activation of AMPA receptors with fast kinetics at a holding potential of -72 mV. The amplitude distribution analysis of the EPSCs indicates that, at both stages, pyramidal-FS connections consisted of multiple functional release sites. The apparent quantal size obtained by decreasing the external calcium ([Ca2+]e) varied from 11 to 29 pA near resting membrane potential. In young rats, pairs of presynaptic action potentials elicited unitary synaptic responses that displayed paired-pulse depression at all tested frequencies. In older animals, inputs from different pyramidal cells onto the same FS interneuron had different paired-pulse response characteristics and, at most of these connections, a switch from depression to facilitation occurred when decreasing the rate of presynaptic stimulation. The balance between facilitation and depression endows pyramidal-FS connections from 5-week-old animals with wide integrative capabilities and confers unique functional properties to each synapse.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous