

Molecular dynamics of the sodium channel pore vary with gating: interactions between P-segment motions and inactivation
- PMID: 10024345
- PMCID: PMC6782169
- DOI: 10.1523/JNEUROSCI.19-05-01577.1999
Molecular dynamics of the sodium channel pore vary with gating: interactions between P-segment motions and inactivation
Abstract
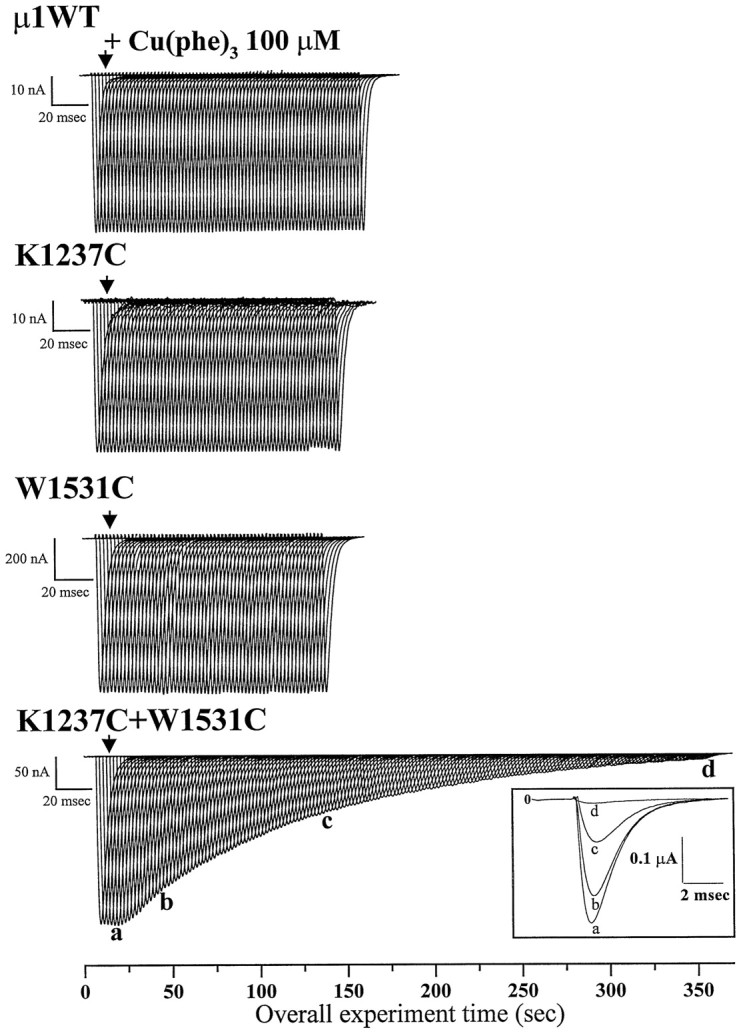
Disulfide trapping studies have revealed that the pore-lining (P) segments of voltage-dependent sodium channels undergo sizable motions on a subsecond time scale. Such motions of the pore may be necessary for selective ion translocation. Although traditionally viewed as separable properties, gating and permeation are now known to interact extensively in various classes of channels. We have investigated the interaction of pore motions and voltage-dependent gating in micro1 sodium channels engineered to contain two cysteines within the P segments. Rates of catalyzed internal disulfide formation (kSS) were measured in K1237C+W1531C mutant channels expressed in oocytes. During repetitive voltage-clamp depolarizations, increasing the pulse duration had biphasic effects on the kSS, which first increased to a maximum at 200 msec and then decreased with longer depolarizations. This result suggested that occupancy of an intermediate inactivation state (IM) facilitates pore motions. Consistent with the known antagonism between alkali metals and a component of slow inactivation, kSS varied inversely with external [Na+]o. We examined the converse relationship, namely the effect of pore flexibility on gating, by measuring recovery from inactivation in Y401C+E758C (YC/EC) channels. Under oxidative conditions, recovery from inactivation was slower than in a reduced environment in which the spontaneous YC/EC cross-link is disrupted. The most prominent effects were slowing of a component with intermediate recovery kinetics, with diminution of its relative amplitude. We conclude that occupancy of an intermediate inactivation state facilitates motions of the P segments; conversely, flexibility of the P segments alters an intermediate component of inactivation.
Figures







References
-
- Bénitah J-P, Balser JR, Marban E, Tomaselli GF. Proton inhibition of sodium channels: mechanism of gating shifts and reduced conductance. J Membr Biol. 1997b;155:121–131. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources