

Vibrio parahaemolyticus thermostable direct hemolysin modulates cytoskeletal organization and calcium homeostasis in intestinal cultured cells
- PMID: 10024554
- PMCID: PMC96440
- DOI: 10.1128/IAI.67.3.1139-1148.1999
Vibrio parahaemolyticus thermostable direct hemolysin modulates cytoskeletal organization and calcium homeostasis in intestinal cultured cells
Abstract
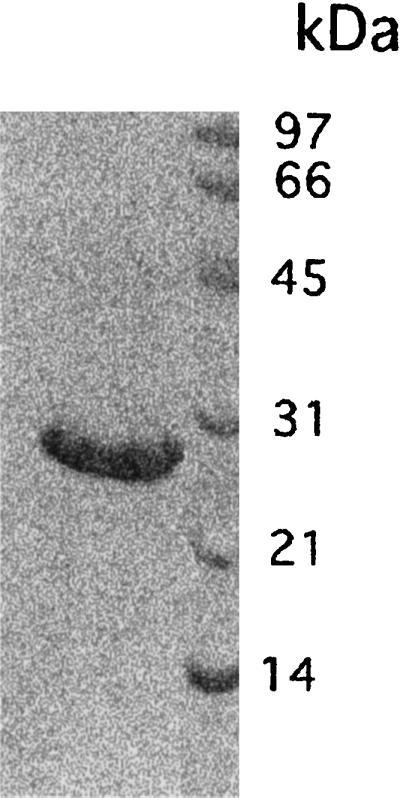
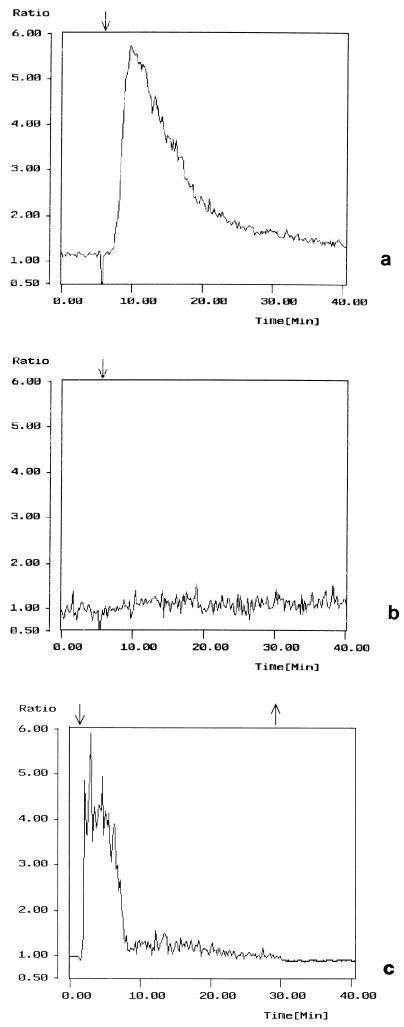
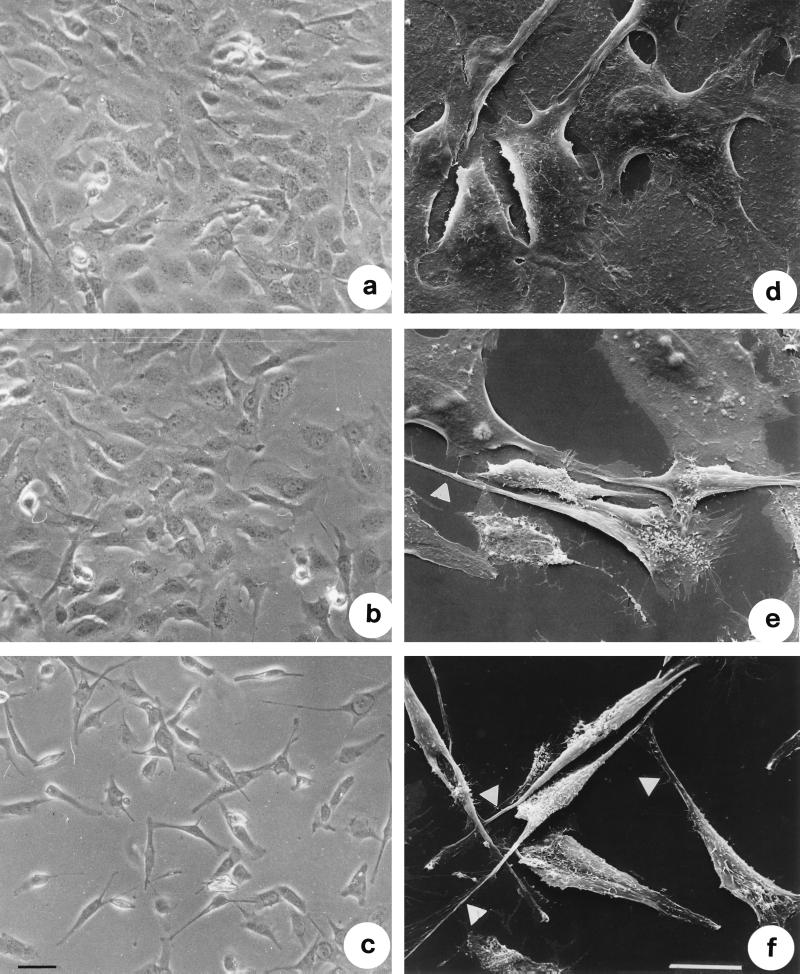
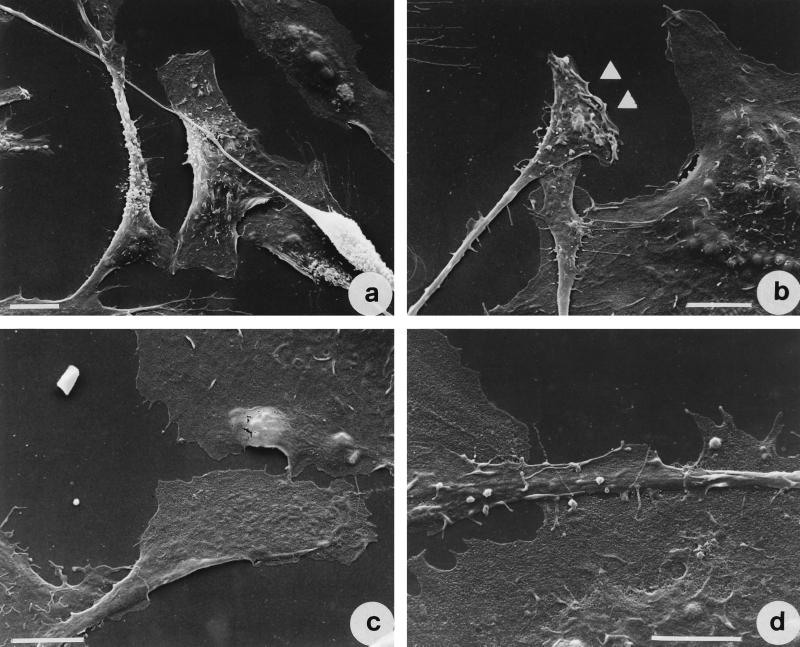
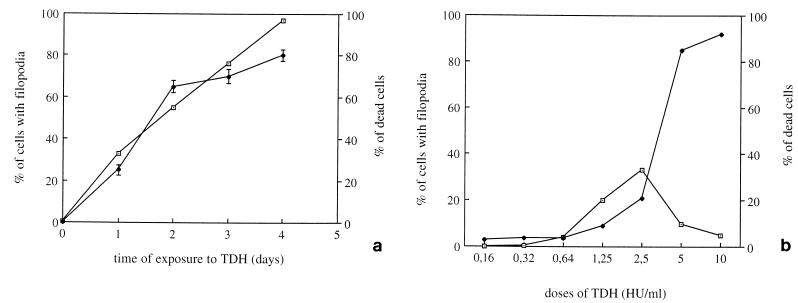
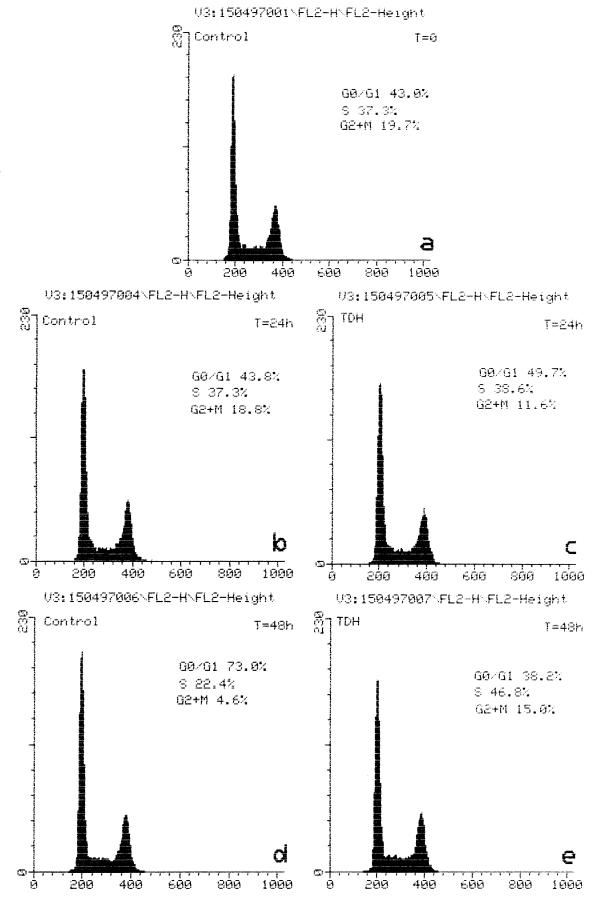
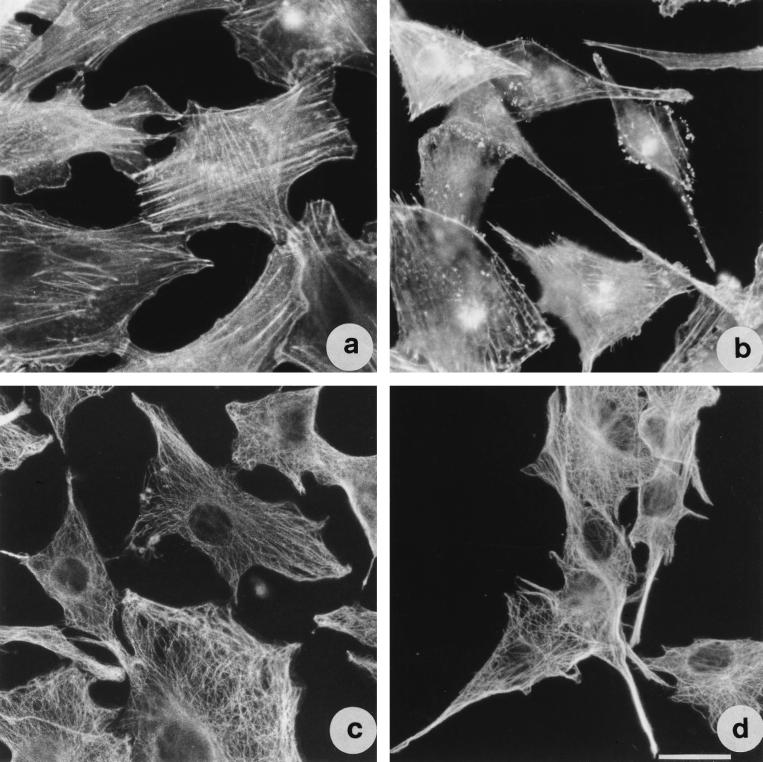
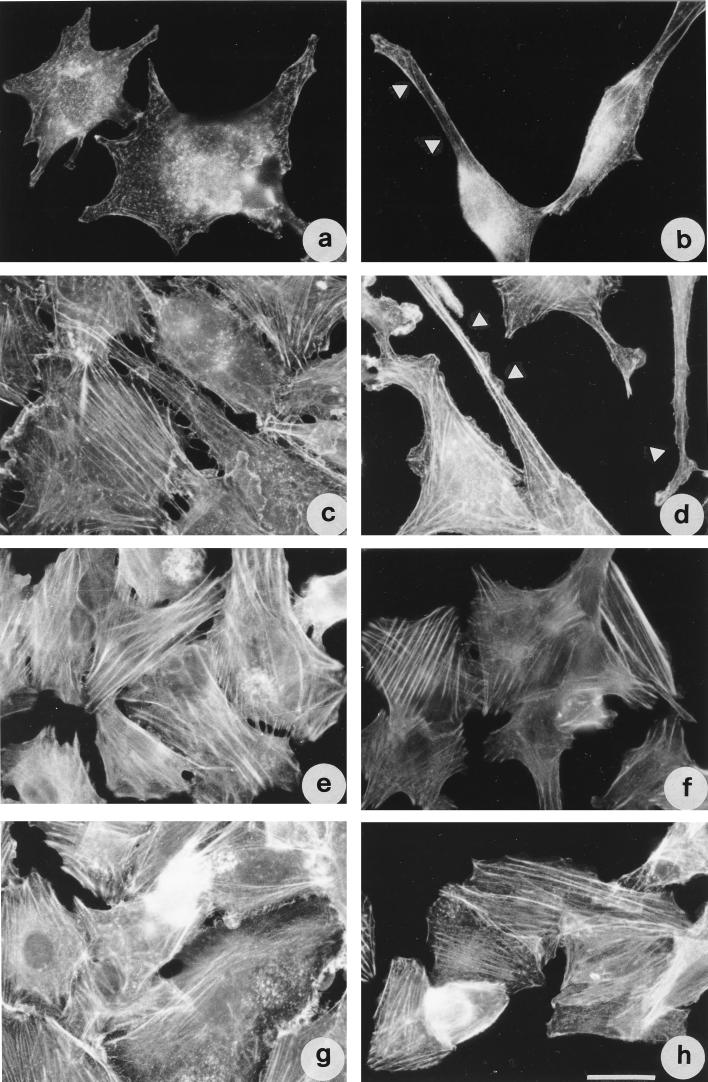
Vibrio parahaemolyticus is a marine bacterium known to be the leading cause of seafood gastroenteritis worldwide. A 46-kDa homodimer protein secreted by this microorganism, the thermostable direct hemolysin (TDH), is considered a major virulence factor involved in bacterial pathogenesis since a high percentage of strains of clinical origin are positive for TDH production. TDH is a pore-forming toxin, and its most extensively studied effect is the ability to cause hemolysis of erythrocytes from different mammalian species. Moreover, TDH induces in a variety of cells cytotoxic effects consisting mainly of cell degeneration which often leads to loss of viability. In this work, we examined the cellular changes induced by TDH in monolayers of IEC-6 cells (derived from the rat crypt small intestine), which represent a useful cell model for studying toxins from enteric bacteria. In experimental conditions allowing cell survival, TDH induces a rapid transient increase in intracellular calcium as well as a significant though reversible decreased rate of progression through the cell cycle. The morphological changes seem to be dependent on the organization of the microtubular network, which appears to be the preferential cytoskeletal element involved in the cellular response to the toxin.
Figures
References
-
- Boquet P, Munro P, Fiorentini C, Just I. Toxins from anaerobic bacteria: specificity and molecular mechanisms of action. Curr Opin Microbiol. 1998;1:66–74. - PubMed
-
- Budd M, Senderowicz A, Sausville E, Duncan K, Korn E. Jasplakinolide, a cytotoxic natural product, induces action polymerization and competitively inhibits the binding of phalloidin to F-actin. J Biol Chem. 1994;269:14869–14871. - PubMed
-
- Cherwonogrodzky J W, Clark A G. The purification of the Kanagawa haemolysin from Vibrio parahaemolyticus. FEMS Microbiol Lett. 1982;15:175. . (Abstract.)
-
- Darzynkiewicz Z, Juan G, Li X, Gorczyca W, Murakami T, Traganos F. Cytometry in cell necrobiology: analysis of apoptosis and accidental cell death (necrosis) Cytometry. 1997;27:1–20. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
