

Reduced differentiation potential of primary MyoD-/- myogenic cells derived from adult skeletal muscle
- PMID: 10037786
- PMCID: PMC2132931
- DOI: 10.1083/jcb.144.4.631
Reduced differentiation potential of primary MyoD-/- myogenic cells derived from adult skeletal muscle
Abstract
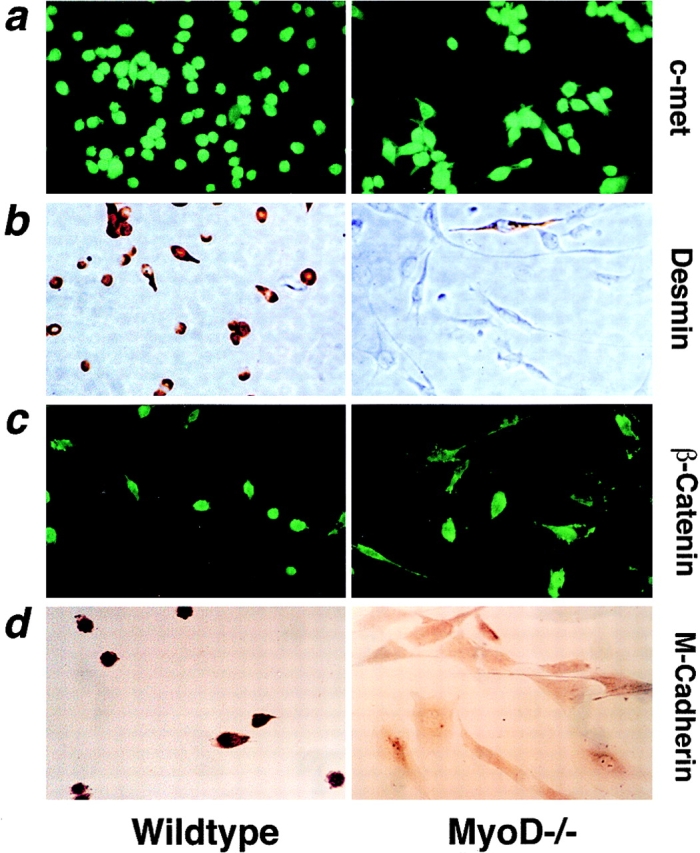
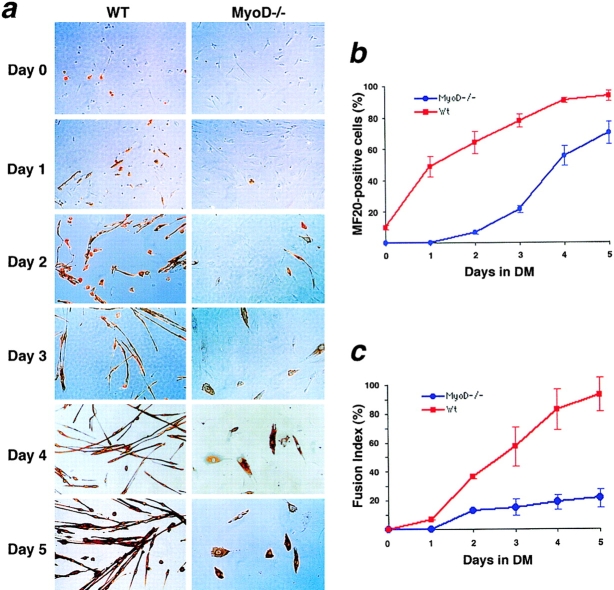
To gain insight into the regeneration deficit of MyoD-/- muscle, we investigated the growth and differentiation of cultured MyoD-/- myogenic cells. Primary MyoD-/- myogenic cells exhibited a stellate morphology distinct from the compact morphology of wild-type myoblasts, and expressed c-met, a receptor tyrosine kinase expressed in satellite cells. However, MyoD-/- myogenic cells did not express desmin, an intermediate filament protein typically expressed in cultured myoblasts in vitro and myogenic precursor cells in vivo. Northern analysis indicated that proliferating MyoD-/- myogenic cells expressed fourfold higher levels of Myf-5 and sixfold higher levels of PEA3, an ETS-domain transcription factor expressed in newly activated satellite cells. Under conditions that normally induce differentiation, MyoD-/- cells continued to proliferate and with delayed kinetics yielded reduced numbers of predominantly mononuclear myocytes. Northern analysis revealed delayed induction of myogenin, MRF4, and other differentiation-specific markers although p21 was upregulated normally. Expression of M-cadherin mRNA was severely decreased whereas expression of IGF-1 was markedly increased in MyoD-/- myogenic cells. Mixing of lacZ-labeled MyoD-/- cells and wild-type myoblasts revealed a strict autonomy in differentiation potential. Transfection of a MyoD-expression cassette restored cytomorphology and rescued the differentiation deficit. We interpret these data to suggest that MyoD-/- myogenic cells represent an intermediate stage between a quiescent satellite cell and a myogenic precursor cell.
Figures







References
-
- Allen RE, Sheehan SM, Taylor RG, Kendall TL, Rice GM. Hepatocyte growth factor activates quiescent skeletal muscle satellite cells in vitro. J Cell Physiol. 1995;165:307–312. - PubMed
-
- Barth AI, Nathke IS, Nelson WJ. Cadherins, catenins and APC protein: interplay between cytoskeletal complexes and signaling pathways. Curr Opin Cell Biol. 1997;9:683–690. - PubMed
-
- Bischoff, R. 1994. The satellite cell and muscle regeneration. In Myogenesis. Vol. 2. A.G. Engel and C. Franszini-Armstrong, editors. McGraw-Hill Inc., New York. 97–118.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
