

ATP dependence of the ICl,swell channel varies with rate of cell swelling. Evidence for two modes of channel activation
- PMID: 10051519
- PMCID: PMC2222898
- DOI: 10.1085/jgp.113.3.441
ATP dependence of the ICl,swell channel varies with rate of cell swelling. Evidence for two modes of channel activation
Abstract
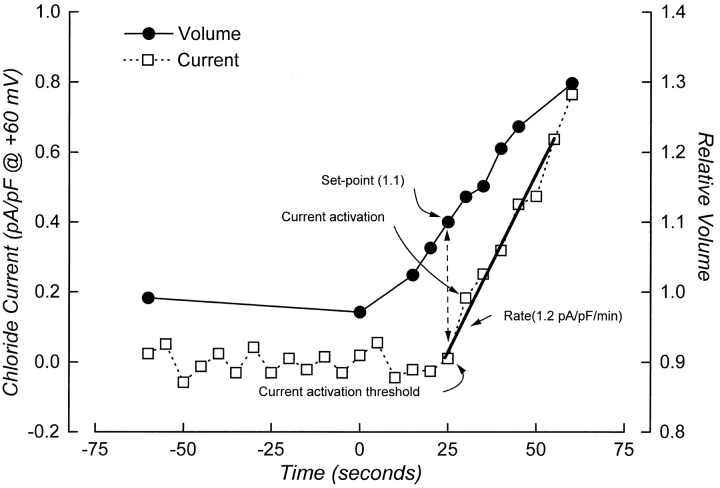
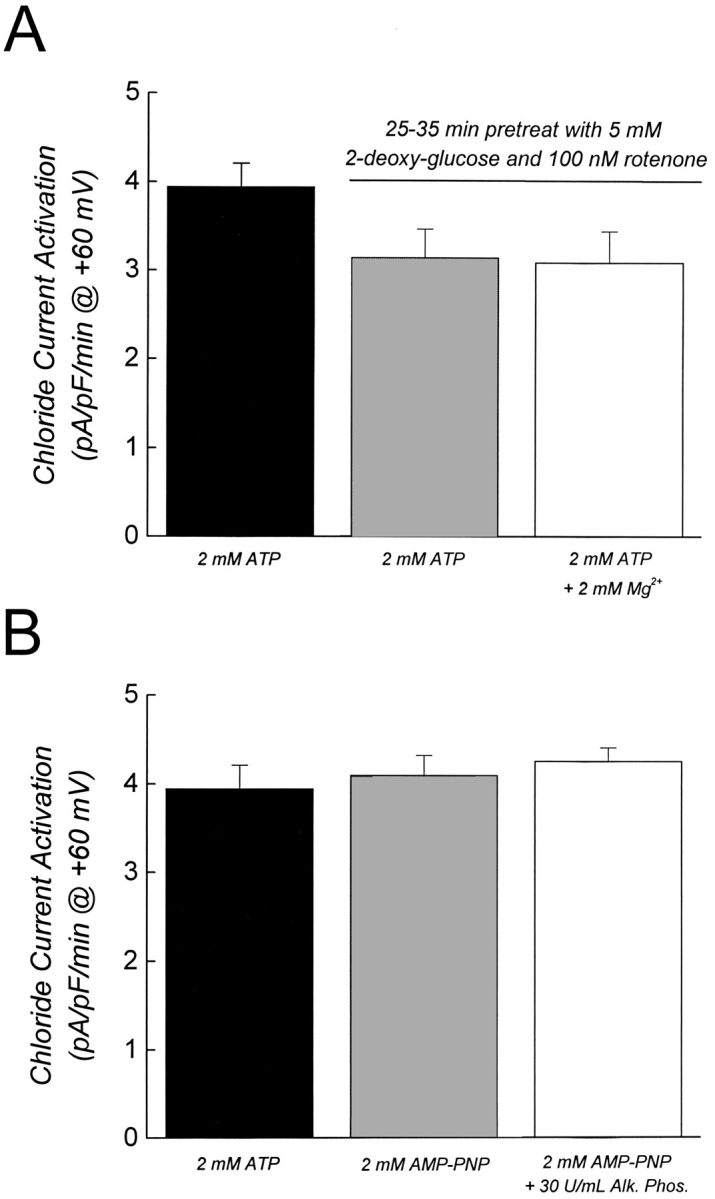
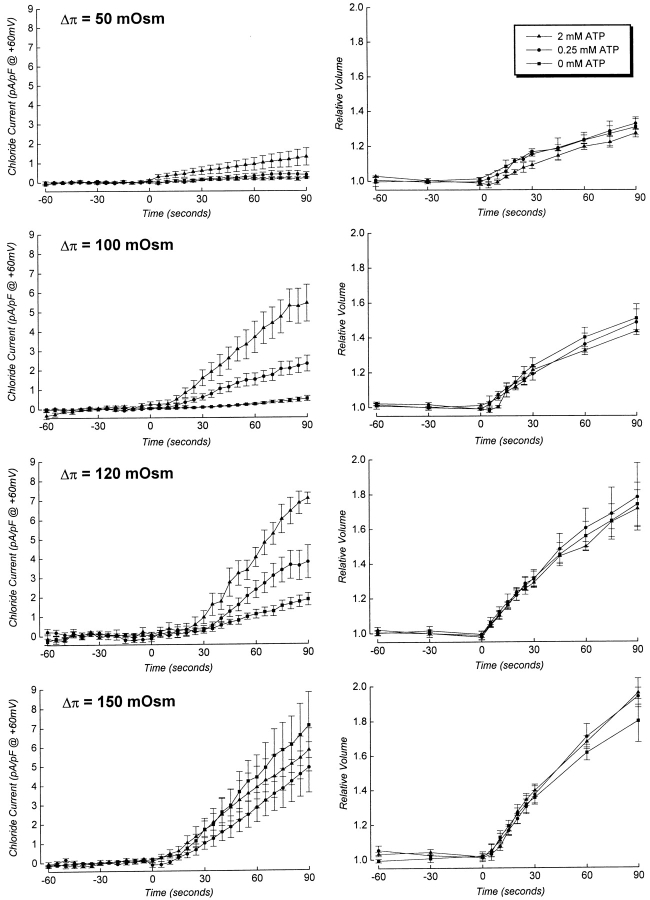
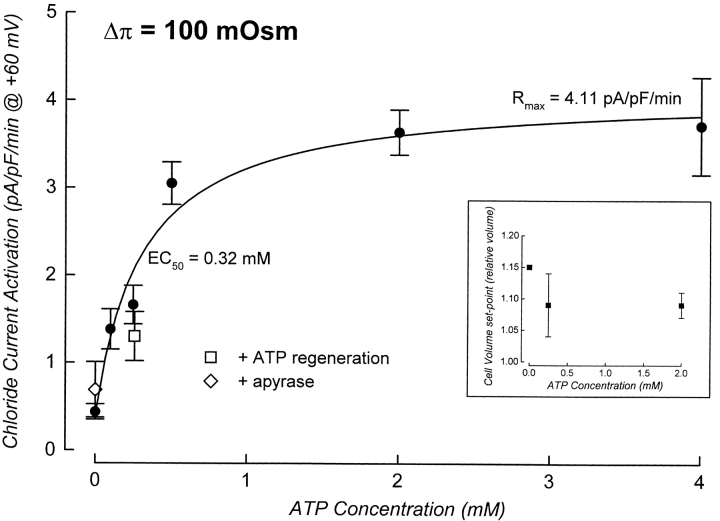
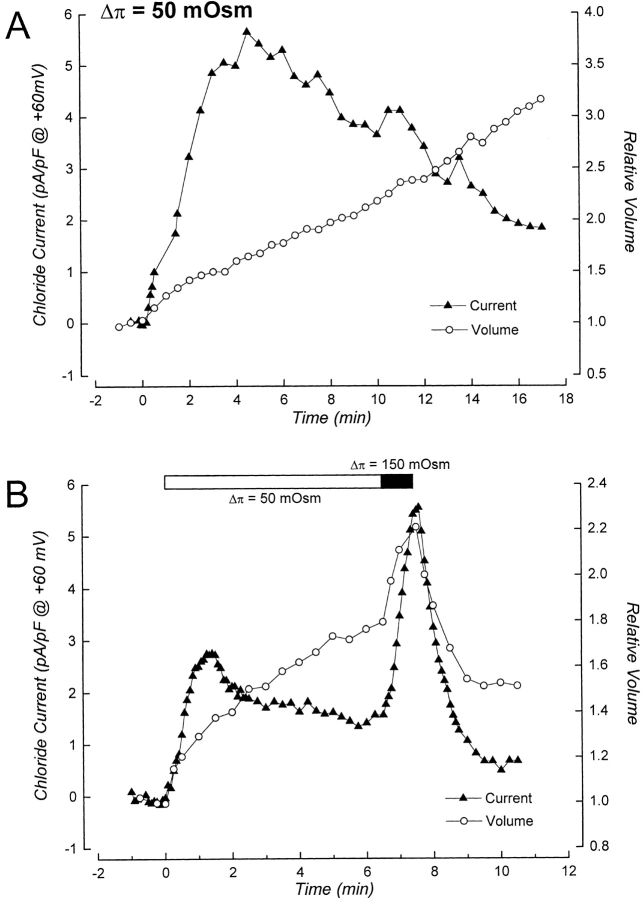
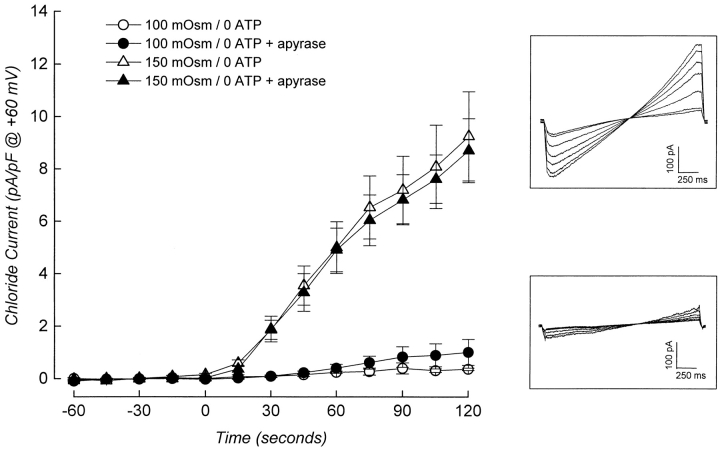
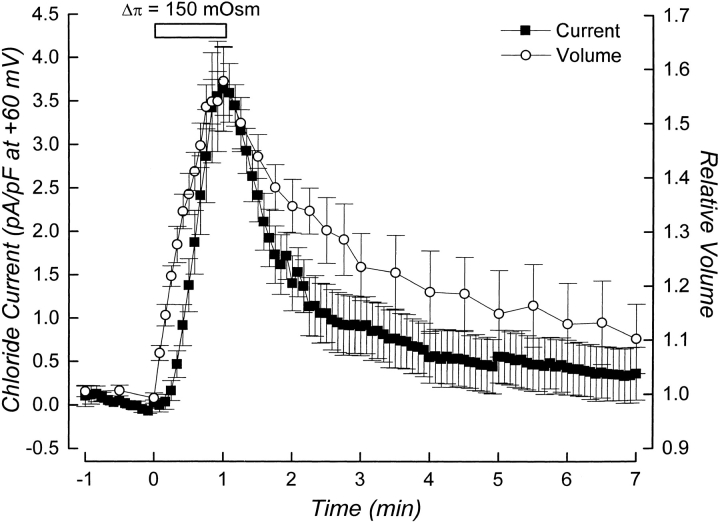
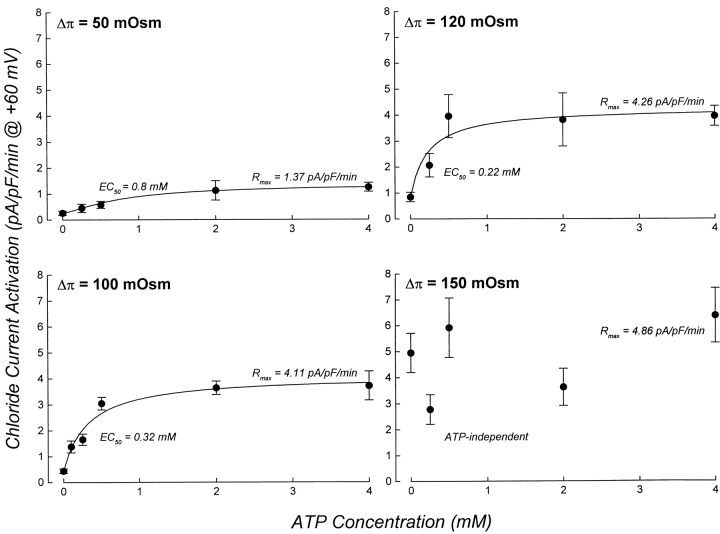
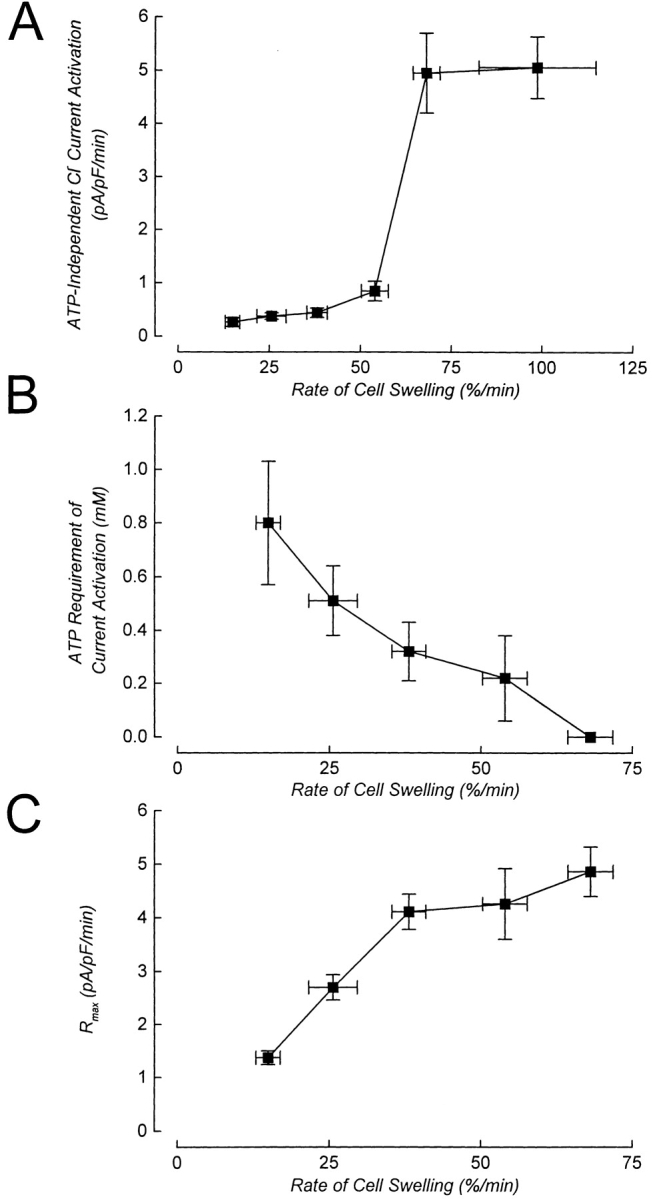
Swelling-induced activation of the outwardly rectifying anion current, ICl, swell, is modulated by intracellular ATP. The mechanisms by which ATP controls channel activation, however, are unknown. Whole cell patch clamp was employed to begin addressing this issue. Endogenous ATP production was inhibited by dialyzing N1E115 neuroblastoma cells for 4-5 min with solutions containing (microM): 40 oligomycin, 5 iodoacetate, and 20 rotenone. The effect of ATP on current activation was observed in the absence of intracellular Mg2+, in cells exposed to extracellular metabolic inhibitors for 25-35 min followed by intracellular dialysis with oligomycin, iodoacetate, and rotenone, after substitution of ATP with the nonhydrolyzable analogue AMP-PNP, and in the presence of AMP-PNP and alkaline phosphatase to dephosphorylate intracellular proteins. These results demonstrate that the ATP dependence of the channel requires ATP binding rather than hydrolysis and/or phosphorylation reactions. When cells were swollen at 15-55%/min in the absence of intracellular ATP, current activation was slow (0.3-0.8 pA/pF per min). ATP concentration increased the rate of current activation up to maximal values of 4-6 pA/pF per min, but had no effect on the sensitivity of the channel to cell swelling. Rate of current activation was a saturable, hyperbolic function of ATP concentration. The EC50 for ATP varied inversely with the rate of cell swelling. Activation of current was rapid (4-6 pA/pF per min) in the absence of ATP when cells were swollen at rates >/=65%/min. Intracellular ATP concentration had no effect on current activation induced by high rates of swelling. Current activation was transient when endogenous ATP was dialyzed out of the cytoplasm of cells swollen at 15%/min. Rundown of the current was reversed by increasing the rate of swelling to 65%/min. These results indicate that the channel and/or associated regulatory proteins are capable of sensing the rate of cell volume increase. We suggest that channel activation occurs via ATP-dependent and -independent mechanisms. Increasing the rate of cell swelling appears to increase the proportion of channels activating via the ATP-independent pathway. These findings have important physiological implications for understanding ICl, swell regulation, the mechanisms by which cells sense volume changes, and volume homeostasis under conditions where cell metabolism is compromised.
Figures
Similar articles
-
Cell swelling activates ATP-dependent voltage-gated chloride channels in M-1 mouse cortical collecting duct cells.J Gen Physiol. 1996 Sep;108(3):177-93. doi: 10.1085/jgp.108.3.177. J Gen Physiol. 1996. PMID: 8882862 Free PMC article.
-
Swelling-induced Cl- current in guinea-pig atrial myocytes: inhibition by glibenclamide.J Physiol. 1997 Nov 15;505 ( Pt 1)(Pt 1):41-52. doi: 10.1111/j.1469-7793.1997.041bc.x. J Physiol. 1997. PMID: 9409470 Free PMC article.
-
Activation of volume-regulated Cl(-) channels by ACh and ATP in Xenopus follicles.J Physiol. 2000 Jun 15;525 Pt 3(Pt 3):721-34. doi: 10.1111/j.1469-7793.2000.00721.x. J Physiol. 2000. PMID: 10856124 Free PMC article.
-
Volume expansion-sensing outward-rectifier Cl- channel: fresh start to the molecular identity and volume sensor.Am J Physiol. 1997 Sep;273(3 Pt 1):C755-89. doi: 10.1152/ajpcell.1997.273.3.C755. Am J Physiol. 1997. PMID: 9316396 Review.
-
Mechanisms of cellular synchronization in the vascular wall. Mechanisms of vasomotion.Dan Med Bull. 2010 Oct;57(10):B4191. Dan Med Bull. 2010. PMID: 21040688 Review.
Cited by
-
Volume-activated chloride currents in fetal human nasopharyngeal epithelial cells.J Membr Biol. 2012 Feb;245(2):107-15. doi: 10.1007/s00232-012-9419-5. Epub 2012 Feb 21. J Membr Biol. 2012. PMID: 22349526
-
Role of volume-regulated and calcium-activated anion channels in cell volume homeostasis, cancer and drug resistance.Channels (Austin). 2015;9(6):380-96. doi: 10.1080/19336950.2015.1089007. Epub 2015 Nov 16. Channels (Austin). 2015. PMID: 26569161 Free PMC article. Review.
-
Potentiation of a voltage-gated proton current in acidosis-induced swelling of rat microglia.J Neurosci. 2000 Oct 1;20(19):7220-7. doi: 10.1523/JNEUROSCI.20-19-07220.2000. J Neurosci. 2000. PMID: 11007878 Free PMC article.
-
LRRC8A-containing chloride channel is crucial for cell volume recovery and survival under hypertonic conditions.Proc Natl Acad Sci U S A. 2021 Jun 8;118(23):e2025013118. doi: 10.1073/pnas.2025013118. Proc Natl Acad Sci U S A. 2021. PMID: 34083438 Free PMC article.
-
Zinc pyrithione activates the volume-regulated anion channel through an antioxidant-sensitive mechanism.Am J Physiol Cell Physiol. 2021 Jun 1;320(6):C1088-C1098. doi: 10.1152/ajpcell.00070.2021. Epub 2021 Apr 7. Am J Physiol Cell Physiol. 2021. PMID: 33826406 Free PMC article.
References
-
- Boese SH, Kinne RHK, Wehner F. Single-channel properties of swelling-activated anion conductance in rat inner medullary collecting duct cells. Am J Physiol. 1996;271:F1224–F1233. - PubMed
-
- Bryan J, Aguilar-Bryan L. The ABCs of ATP-sensitive potassium channels: more pieces of the puzzle. Curr Opin Cell Biol. 1997;9:553–559. - PubMed
-
- Cannon CL, Basavappa S, Strange K. Intracellular ionic strength regulates the volume sensitivity of a swelling-activated anion channel. Am J Physiol. 1998;275:C416–C422. - PubMed
-
- Chicurel ME, Chen CS, Ingber DE. Cellular control lies in the balance of forces. Curr Opin Cell Biol. 1998;10:232–239. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
