

Blockade of N-methyl-D-aspartate receptor activation suppresses learning-induced synaptic elimination
- PMID: 10051669
- PMCID: PMC26811
- DOI: 10.1073/pnas.96.5.2485
Blockade of N-methyl-D-aspartate receptor activation suppresses learning-induced synaptic elimination
Abstract
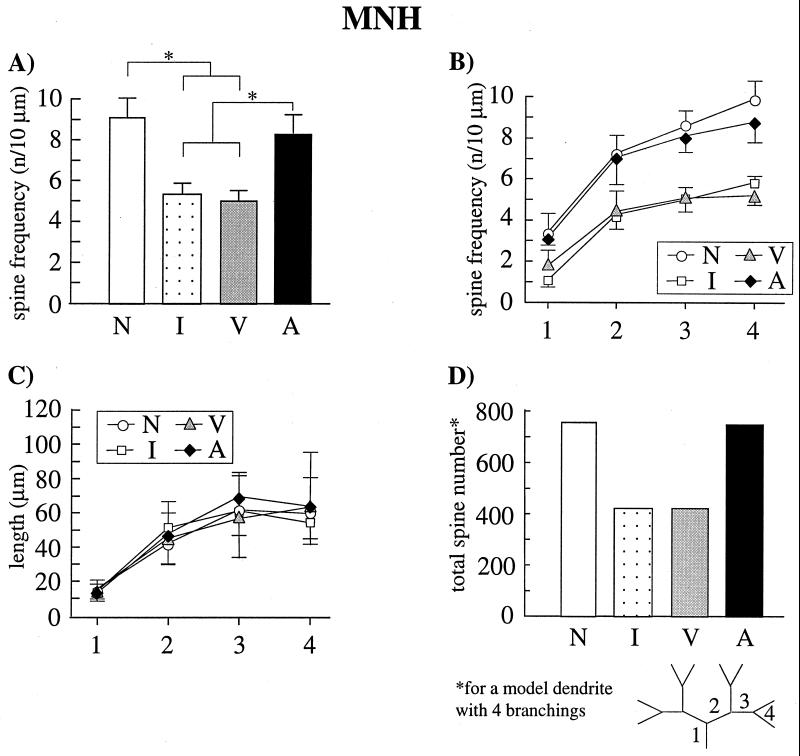
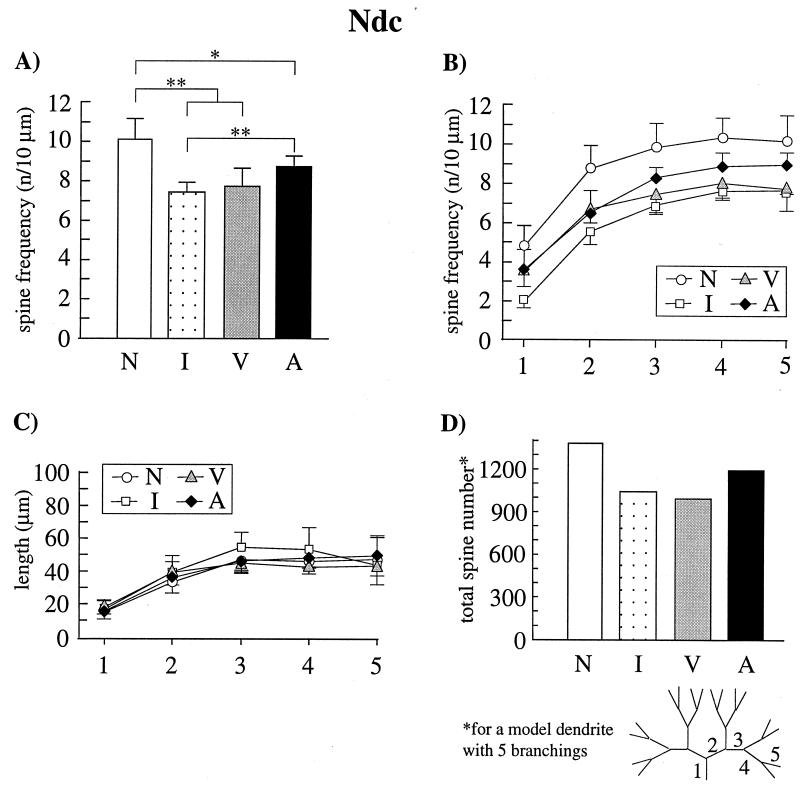
Auditory filial imprinting in the domestic chicken is accompanied by a dramatic loss of spine synapses in two higher associative forebrain areas, the mediorostral neostriatum/hyperstriatum ventrale (MNH) and the dorsocaudal neostriatum (Ndc). The cellular mechanisms that underlie this learning-induced synaptic reorganization are unclear. We found that local pharmacological blockade of N-methyl-D-aspartate (NMDA) receptors in the MNH, a manipulation that has been shown previously to impair auditory imprinting, suppresses the learning-induced spine reduction in this region. Chicks treated with the NMDA receptor antagonist 2-amino-5-phosphonovaleric acid (APV) during the behavioral training for imprinting (postnatal day 0-2) displayed similar spine frequencies at postnatal day 7 as naive control animals, which, in both groups, were significantly higher than in imprinted animals. Because the average dendritic length did not differ between the experimental groups, the reduced spine frequency can be interpreted as a reduction of the total number of spine synapses per neuron. In the Ndc, which is reciprocally connected with the MNH and not directly influenced by the injected drug, learning-induced spine elimination was partly suppressed. Spine frequencies of the APV-treated, behaviorally trained but nonimprinted animals were higher than in the imprinted animals but lower than in the naive animals. These results provide evidence that NMDA receptor activation is required for the learning-induced selective reduction of spine synapses, which may serve as a mechanism of information storage specific for juvenile emotional learning events.
Figures


References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
