

Distinct signaling pathways mediate touch and osmosensory responses in a polymodal sensory neuron
- PMID: 10066248
- PMCID: PMC6782580
- DOI: 10.1523/JNEUROSCI.19-06-01952.1999
Distinct signaling pathways mediate touch and osmosensory responses in a polymodal sensory neuron
Abstract
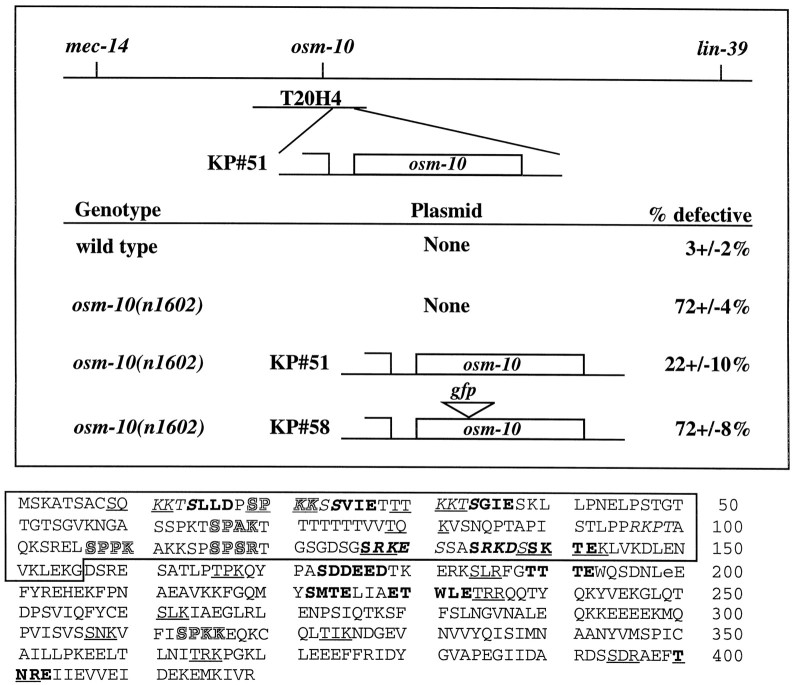
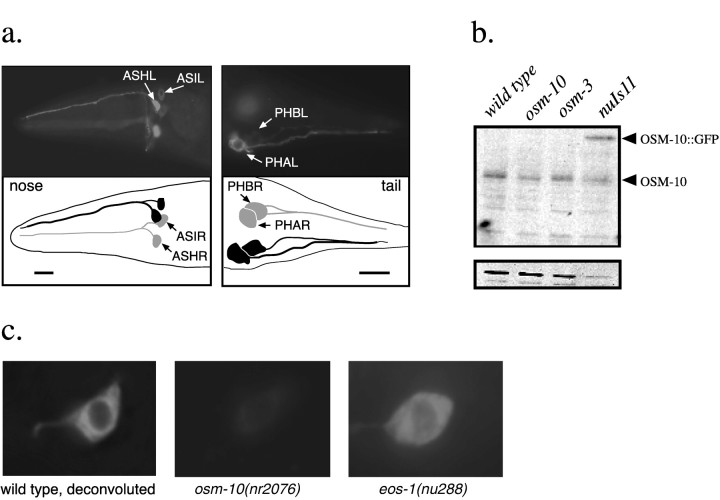
The Caenorhabditis elegans ASH sensory neurons mediate responses to nose touch, hyperosmolarity, and volatile repellent chemicals. We show here that distinct signaling pathways mediate the responses to touch and hyperosmolarity. ASH neurons distinguish between these stimuli because habituation to nose touch has no effect on the response to high osmolarity or volatile chemicals (1-octanol). Mutations in osm-10 eliminate the response to hyperosmolarity but have no effect on responses to nose touch or to volatile repellents. OSM-10 is a novel cytosolic protein expressed in ASH and three other classes of sensory neurons. Mutations in two other osmosensory-defective genes, eos-1 and eos-2, interact genetically with osm-10. Our analysis suggests that nose touch sensitivity and osmosensation occur via distinct signaling pathways in ASH and that OSM-10 is required for osmosensory signaling.
Figures
References
-
- Albert PS, Brown SJ, Riddle DL. Sensory control of dauer larva formation in C. elegans. J Comp Neurol. 1981;198:435–451. - PubMed
-
- Avery L, Horvitz H. Effects of killing identified pharyngeal neurons on feeding behavior in Caenorhabditis elegans. Neuron. 1989;3:473–485. - PubMed
-
- Bargmann CI, Thomas JH, Horvitz HR. Chemosensory cell function in the behavior and development of Caenorhabditis elegans. Cold Spring Harb Symp Quant Biol. 1990;55:529–538. - PubMed
-
- Brewster J, de Valoir T, Dwyer N, Winter E, Gustin M. An osmosensing signal transduction pathway in yeast. Science. 1993;259:1760–1763. - PubMed
-
- Caterina M, Schumacher M, Tominaga M, Rosen T, Levine J, Julius D. The capsaicin receptor: a heat-activated ion channel in the pain pathway. Nature. 1997;389:816. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous