

Leukemia inhibitory factor and ciliary neurotrophic factor cause dendritic retraction in cultured rat sympathetic neurons
- PMID: 10066264
- PMCID: PMC6782564
- DOI: 10.1523/JNEUROSCI.19-06-02113.1999
Leukemia inhibitory factor and ciliary neurotrophic factor cause dendritic retraction in cultured rat sympathetic neurons
Abstract
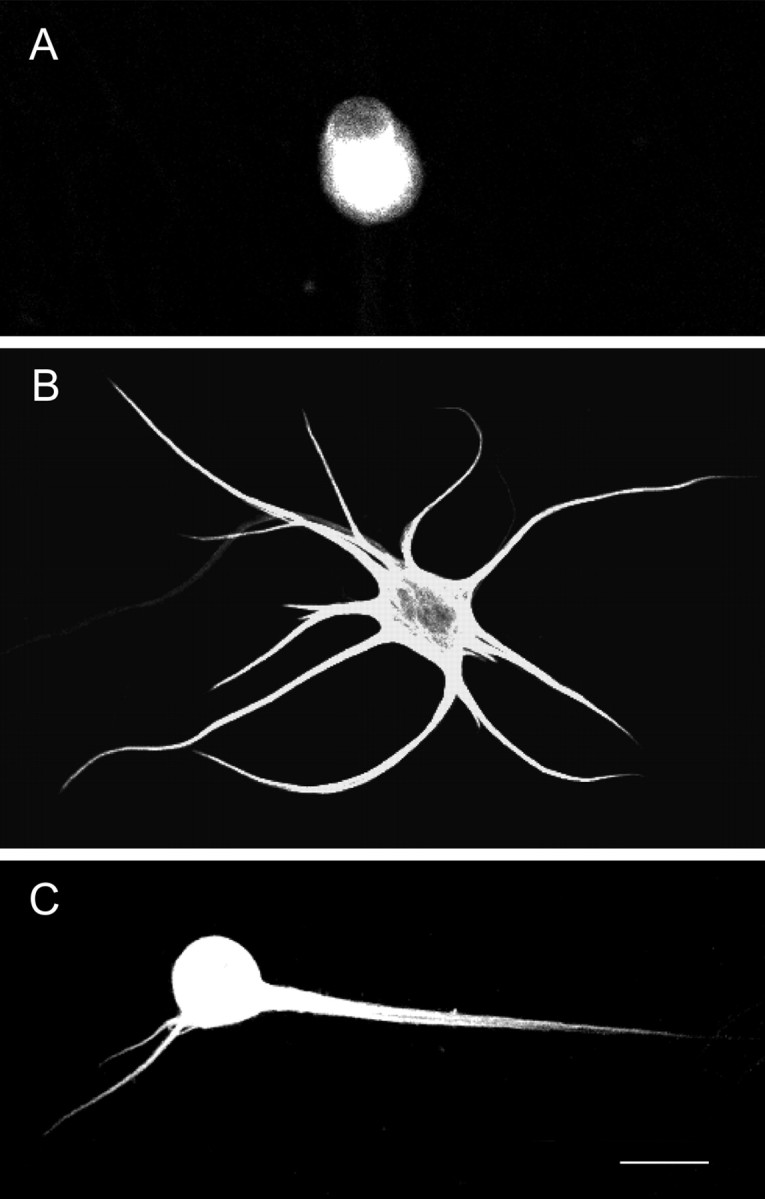
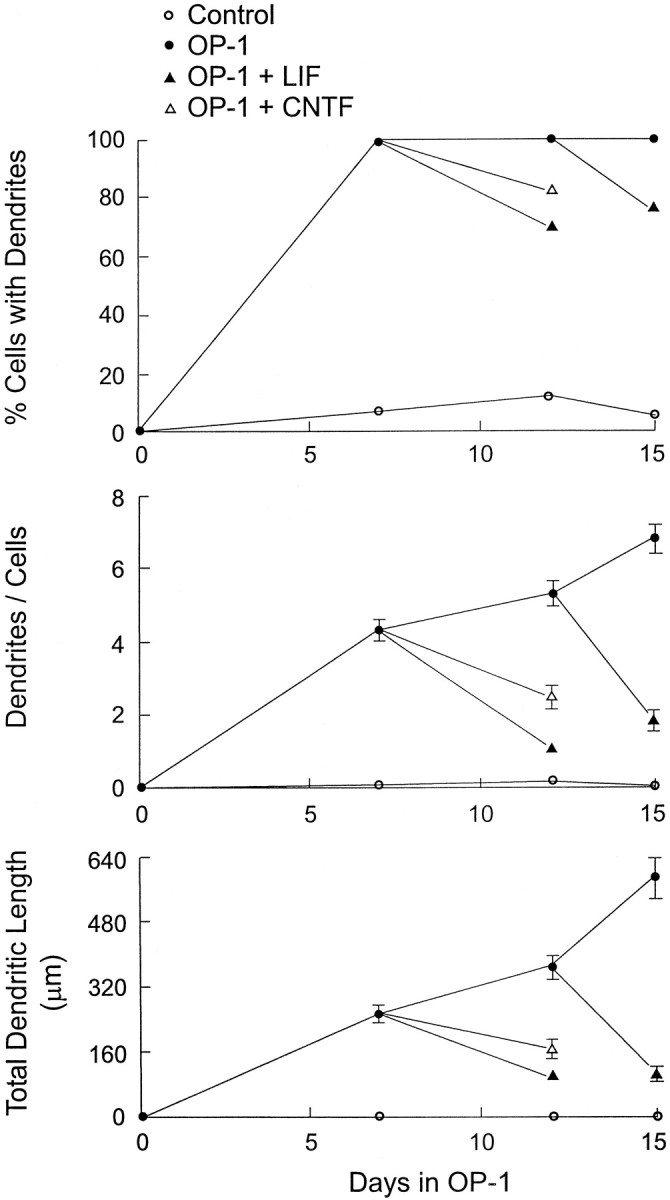
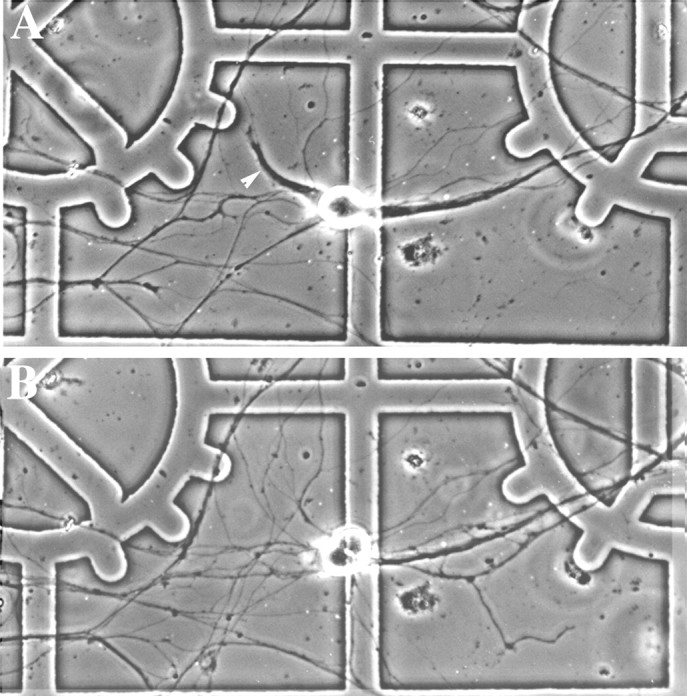
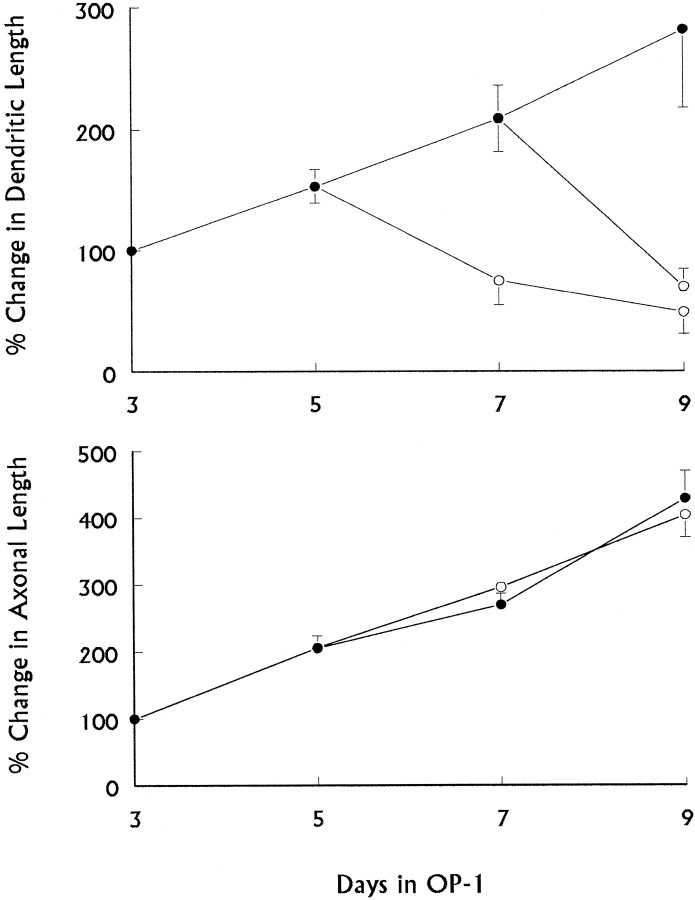
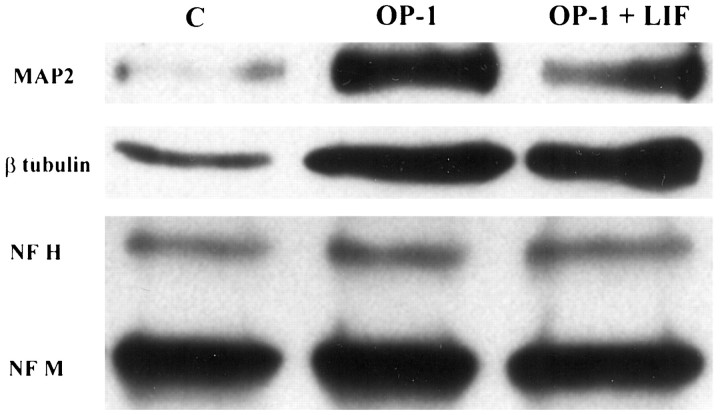
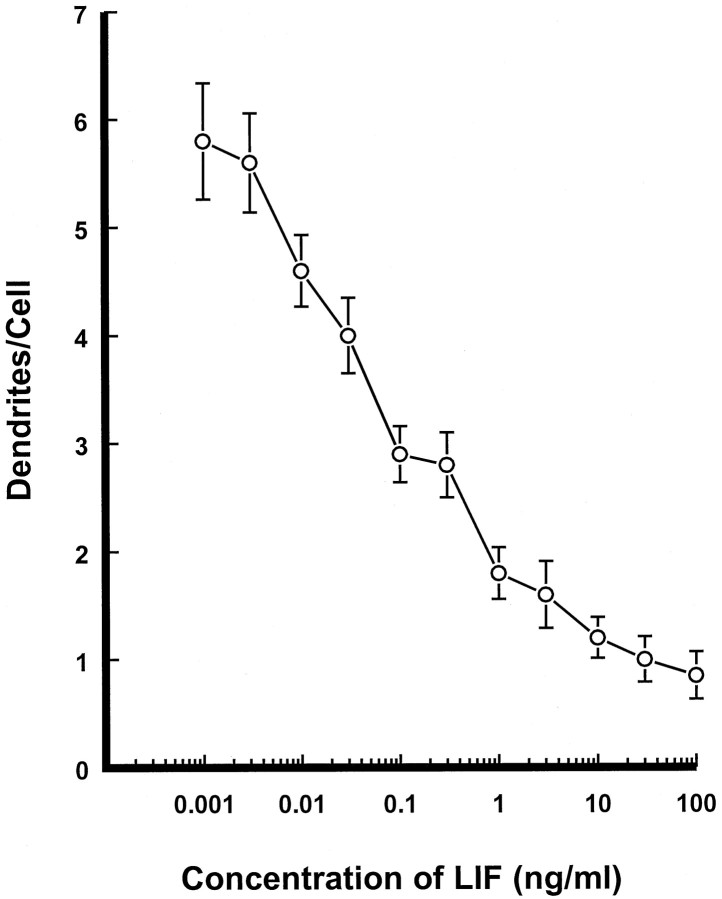
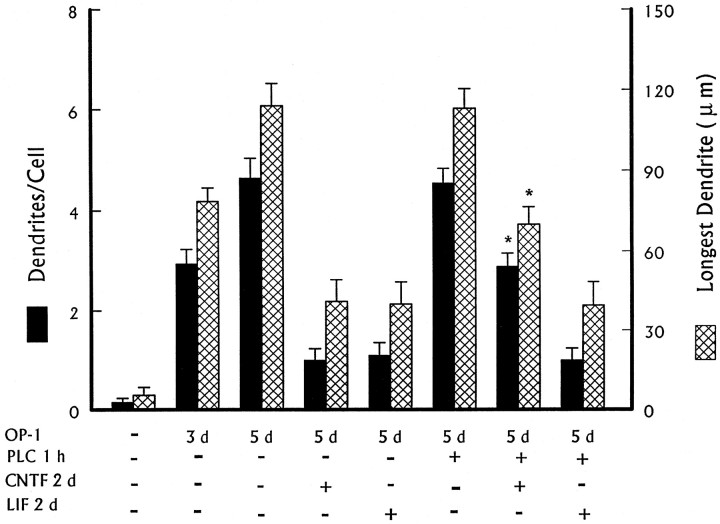
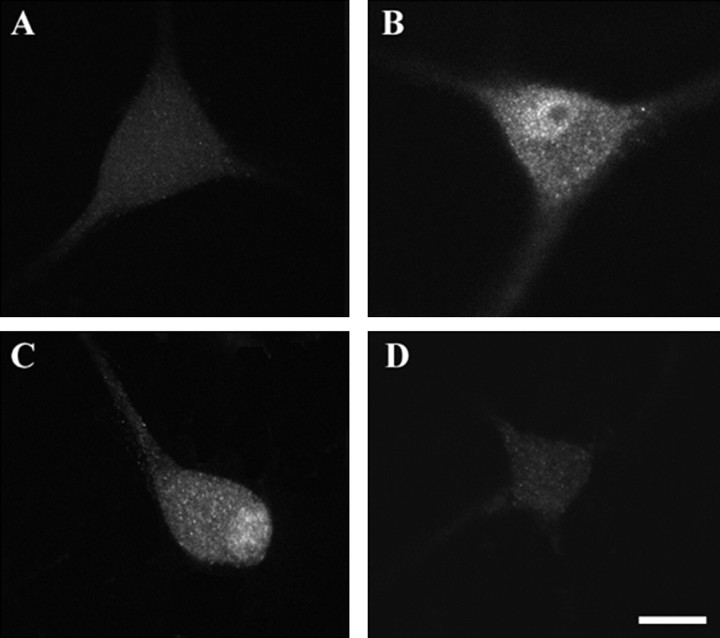
Dendritic retraction occurs in many regions of the developing brain and also after neural injury. However, the molecules that regulate this important regressive process remain largely unknown. Our data indicate that leukemia inhibitory factor (LIF) and ciliary neurotrophic factor (CNTF) cause sympathetic neurons to retract their dendrites in vitro, ultimately leading to an approximately 80% reduction in the size of the arbor. The dendritic retraction induced by LIF exhibited substantial specificity because it was not accompanied by changes in cell number, in the rate of axonal growth, or in the expression of axonal cytoskeletal elements. An antibody to gp130 blocked the effects of LIF and CNTF, and both cytokines induced phosphorylation and nuclear translocation of stat3. Moreover, addition of soluble interleukin-6 (IL-6) receptor to the medium endowed IL-6 with the ability to cause dendritic regression. These data indicate that ligands activating the gp130 pathway have the ability to profoundly alter neuronal cell shape and polarity by selectively causing the retraction of dendrites.
Figures


References
-
- Armengol JA, Sotelo C. Early dendritic development of Purkinje cells in the rat cerebellum. A light and electron microscopic study using axonal tracing in “in vitro” slices. Dev Brain Res. 1991;64:95–114. - PubMed
-
- Bauer J, Strauss S, Schreiter-Gasser U, Ganter U, Schlegel P, Witt I, Yolk B, Berger M. Interleukin-6 and alpha-2 macroglobulin indicate an acute phase state in Alzheimer’s disease cortices. FEBS Lett. 1991;285:111–114. - PubMed
-
- Carlson CD, Bai Y, Ding M, Jonakait GM, Hart RP. Interleukin 1 involvement in the induction of leukemia inhibitory factor mRNA expression following axotomy of sympathetic ganglia. J Neuroimmunol. 1996;70:181–190. - PubMed
-
- Carter BD, Lewin GR. Neurotrophins live or let die: does p75ntr decide? Neuron. 1997;18:187–190. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous