

Mechanisms of arthropod transmission of plant and animal viruses
- PMID: 10066833
- PMCID: PMC98959
- DOI: 10.1128/MMBR.63.1.128-148.1999
Mechanisms of arthropod transmission of plant and animal viruses
Abstract
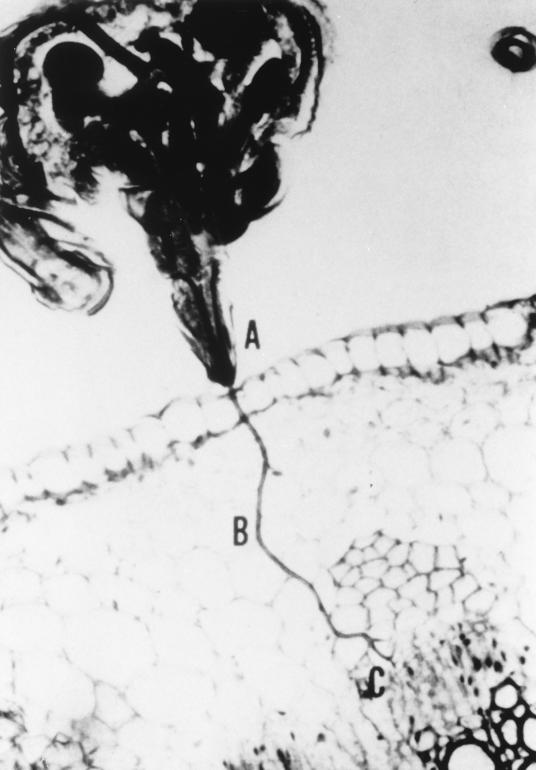
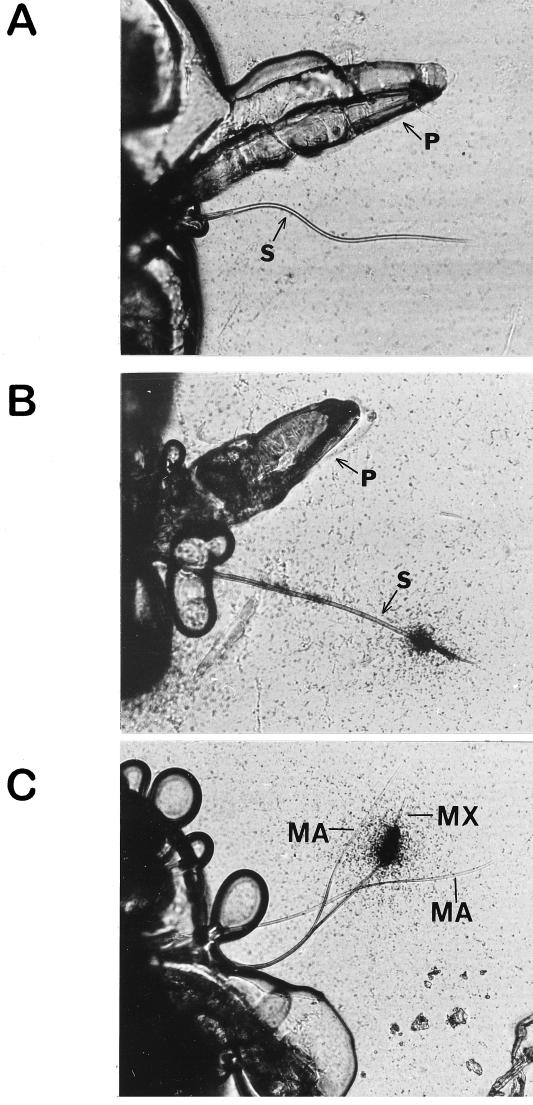
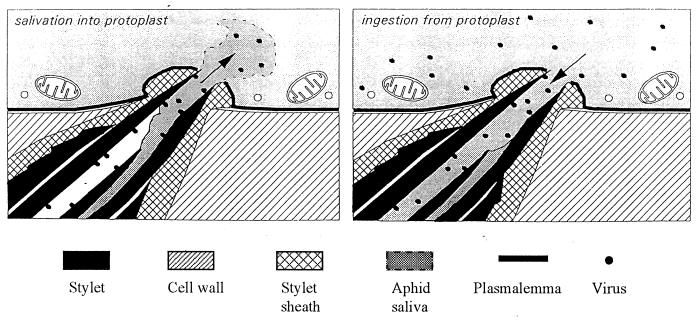
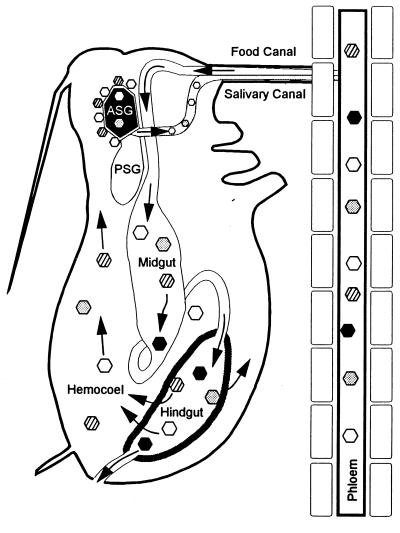
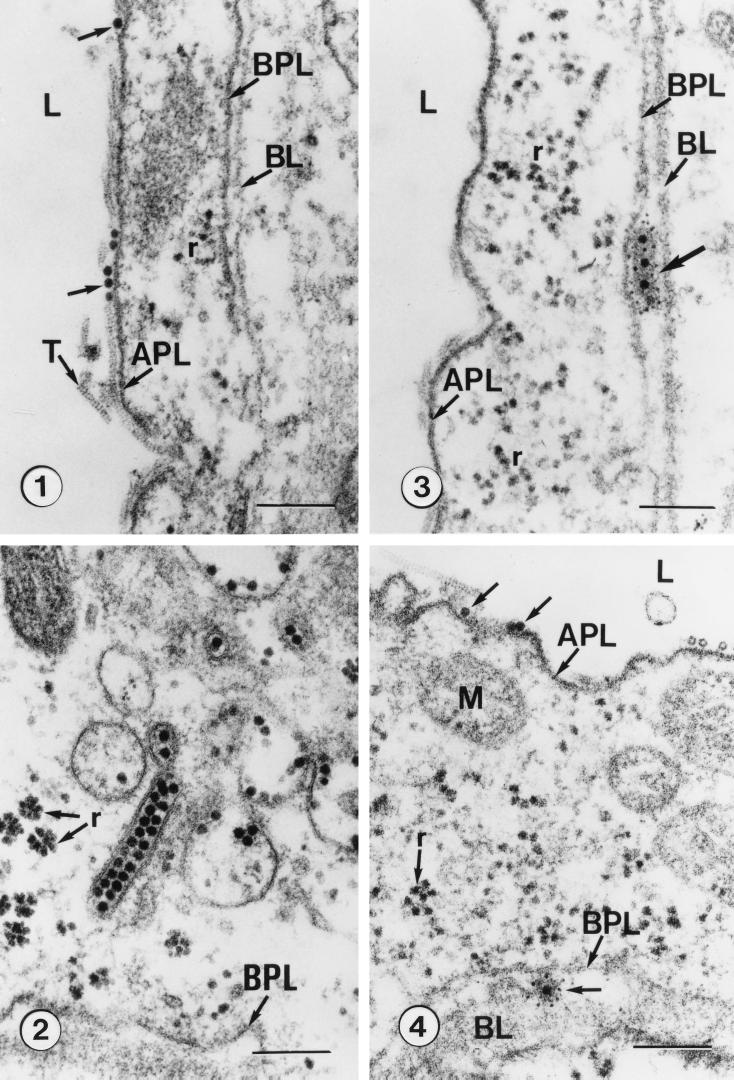
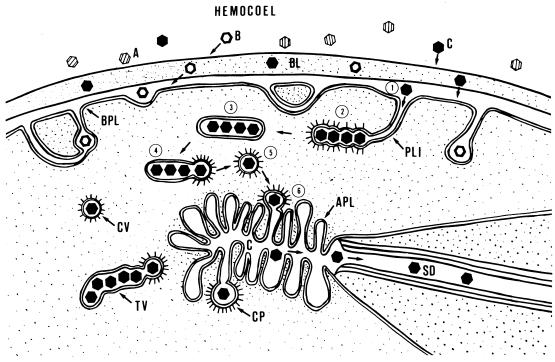
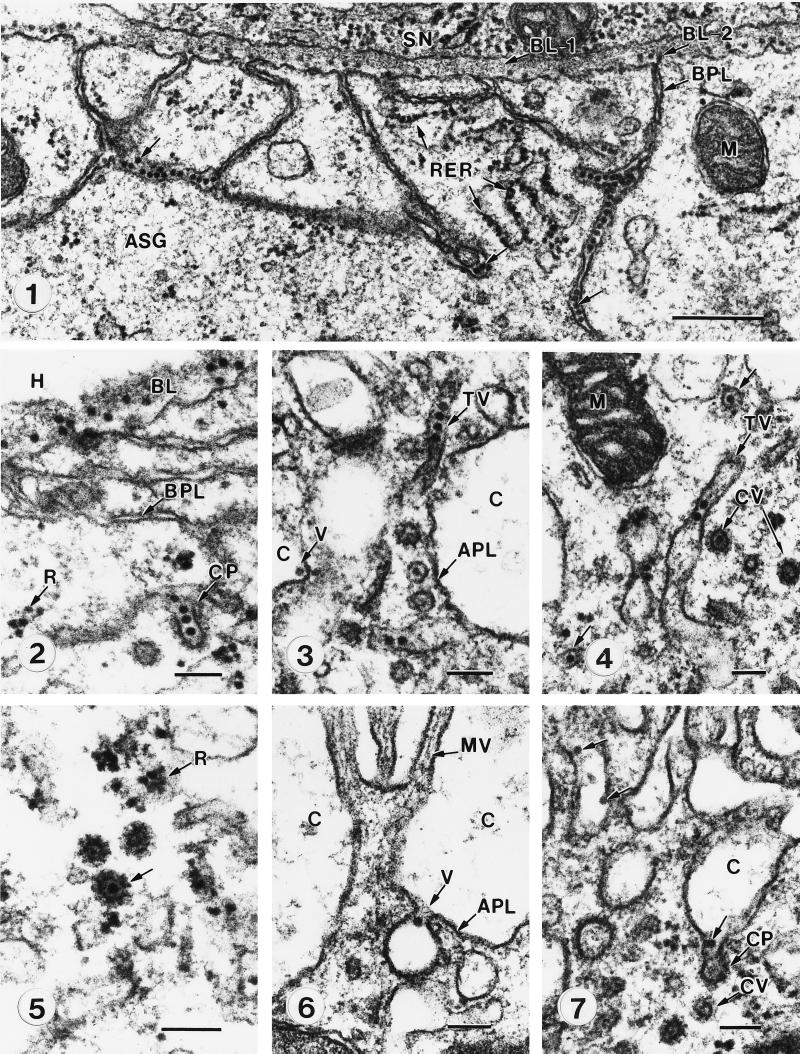
A majority of the plant-infecting viruses and many of the animal-infecting viruses are dependent upon arthropod vectors for transmission between hosts and/or as alternative hosts. The viruses have evolved specific associations with their vectors, and we are beginning to understand the underlying mechanisms that regulate the virus transmission process. A majority of plant viruses are carried on the cuticle lining of a vector's mouthparts or foregut. This initially appeared to be simple mechanical contamination, but it is now known to be a biologically complex interaction between specific virus proteins and as yet unidentified vector cuticle-associated compounds. Numerous other plant viruses and the majority of animal viruses are carried within the body of the vector. These viruses have evolved specific mechanisms to enable them to be transported through multiple tissues and to evade vector defenses. In response, vector species have evolved so that not all individuals within a species are susceptible to virus infection or can serve as a competent vector. Not only are the virus components of the transmission process being identified, but also the genetic and physiological components of the vectors which determine their ability to be used successfully by the virus are being elucidated. The mechanisms of arthropod-virus associations are many and complex, but common themes are beginning to emerge which may allow the development of novel strategies to ultimately control epidemics caused by arthropod-borne viruses.
Figures
References
-
- Al K N, Covey S N. Variation in biological properties of cauliflower mosaic virus clones. J Gen Virol. 1994;75:3137–3145. - PubMed
-
- Ammar E D. Propagative transmission of plant and animal viruses by insects: factors affecting vector specificity and competence. Adv Dis Vector Res. 1994;10:289–331.
-
- Ammar E D, Jarlfors U, Pirone T P. Association of potyvirus helper component protein with virions and the cuticle lining the maxillary food canal and foregut of an aphid vector. Phytopathology. 1994;84:1054–1060.
-
- Backus E A. Anatomical and sensory mechanisms of planthopper and leafhopper feeding behavior. In: Nault L R, Rodriguez J G, editors. The leafhoppers and planthoppers. New York, N.Y: John Wiley & Sons, Inc.; 1985. pp. 163–194.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
