

Kir2.1 encodes the inward rectifier potassium channel in rat arterial smooth muscle cells
- PMID: 10066894
- PMCID: PMC2269194
- DOI: 10.1111/j.1469-7793.1999.639ab.x
Kir2.1 encodes the inward rectifier potassium channel in rat arterial smooth muscle cells
Abstract
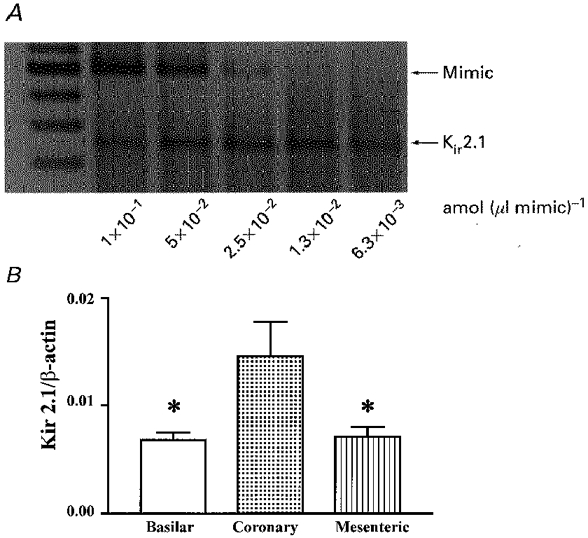
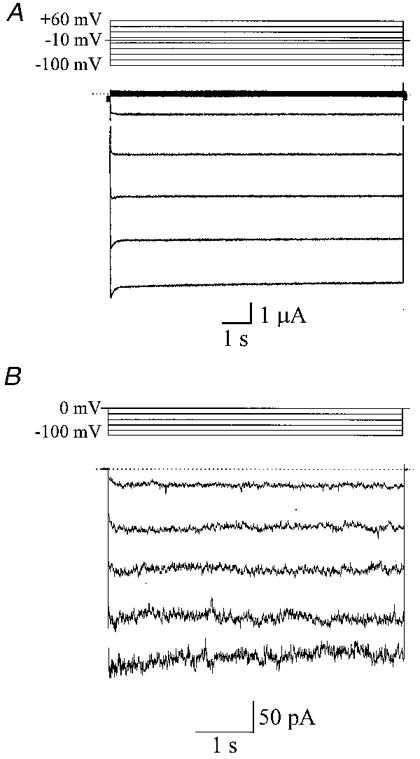
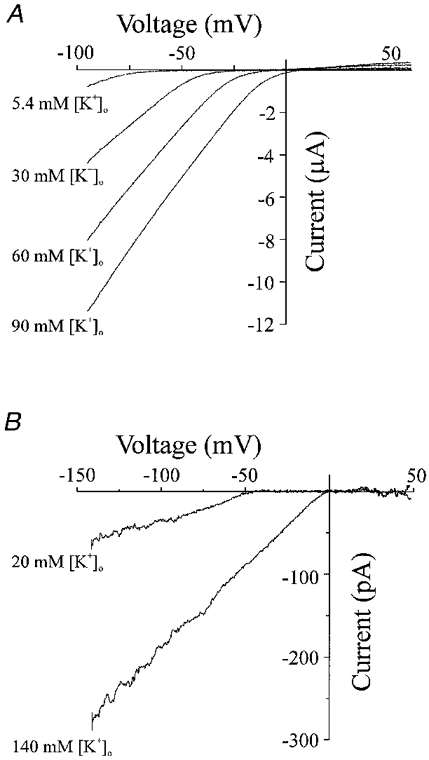
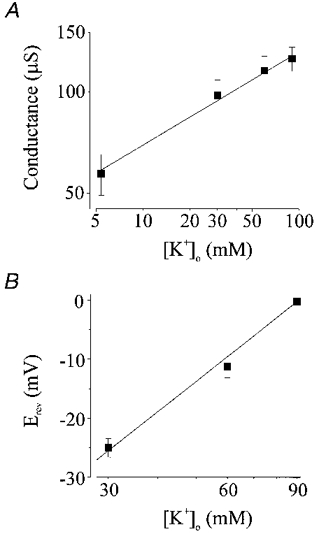
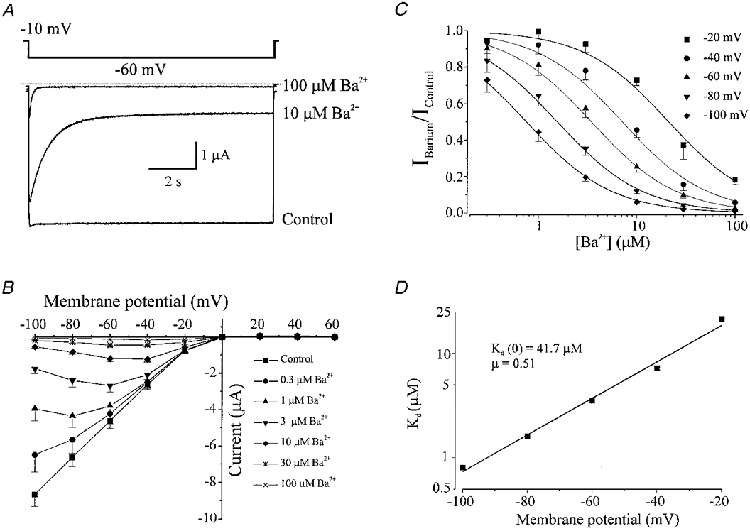
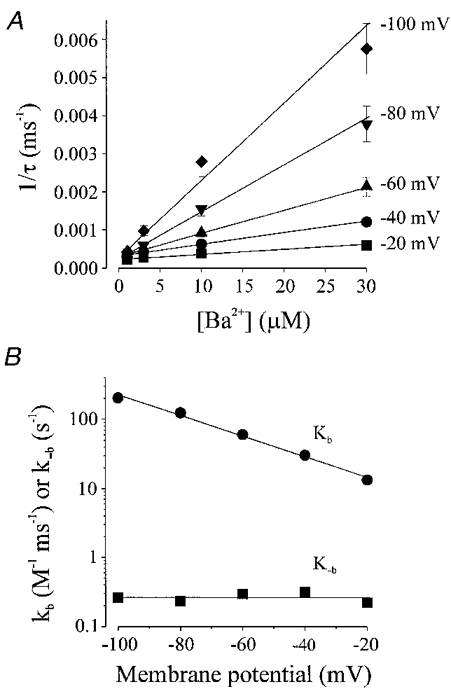
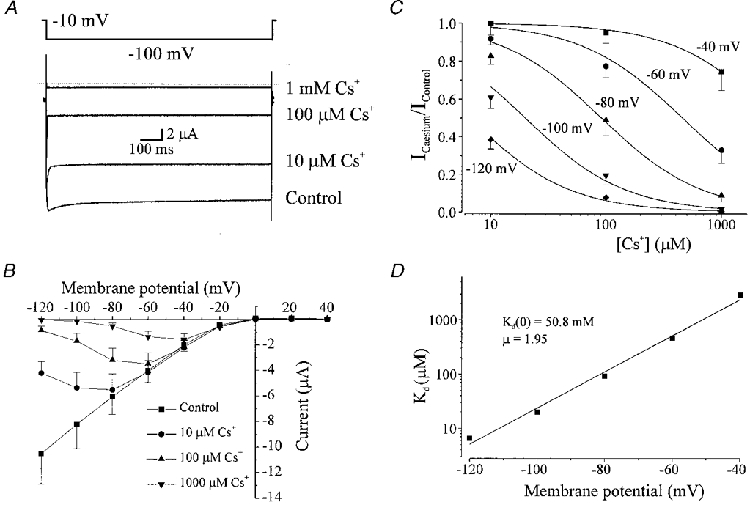
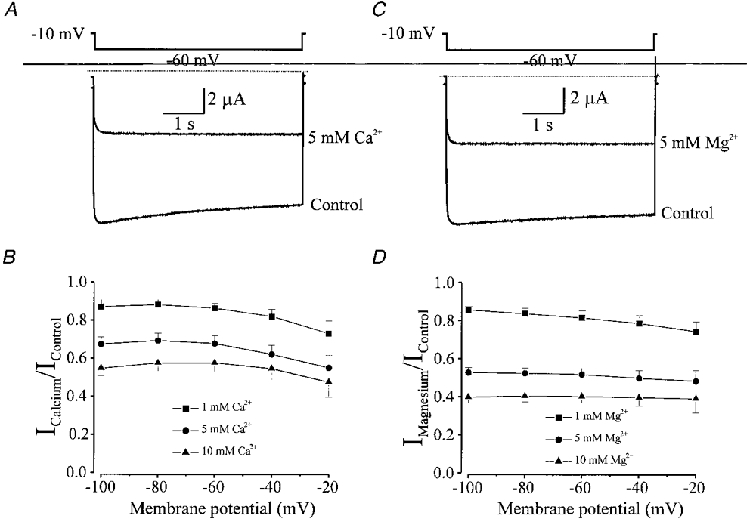
1. The molecular nature of the strong inward rectifier K+ channel in vascular smooth muscle was explored by using isolated cell RT-PCR, cDNA cloning and expression techniques. 2. RT-PCR of RNA from single smooth muscle cells of rat cerebral (basilar), coronary and mesenteric arteries revealed transcripts for Kir2.1. Transcripts for Kir2.2 and Kir2.3 were not found. 3. Quantitative PCR analysis revealed significant differences in transcript levels of Kir2.1 between the different vascular preparations (n = 3; P < 0.05). A two-fold difference was detected between Kir2.1 mRNA and beta-actin mRNA in coronary arteries when compared with relative levels measured in mesenteric and basilar preparations. 4. Kir2.1 was cloned from rat mesenteric vascular smooth muscle cells and expressed in Xenopus oocytes. Currents were strongly inwardly rectifying and selective for K+. 5. The effect of extracellular Ba2+, Ca2+, Mg2+ and Cs2+ ions on cloned Kir2.1 channels expressed in Xenopus oocytes was examined. Ba2+ and Cs+ block were steeply voltage dependent, whereas block by external Ca2+ and Mg2+ exhibited little voltage dependence. The apparent half-block constants and voltage dependences for Ba2+, Cs+, Ca2+ and Mg2+ were very similar for inward rectifier K+ currents from native cells and cloned Kir2.1 channels expressed in oocytes. 6. Molecular studies demonstrate that Kir2.1 is the only member of the Kir2 channel subfamily present in vascular arterial smooth muscle cells. Expression of cloned Kir2.1 in Xenopus oocytes resulted in inward rectifier K+ currents that strongly resemble those that are observed in native vascular arterial smooth muscle cells. We conclude that Kir2.1 encodes for inward rectifier K+ channels in arterial smooth muscle.
Figures

References
-
- Bonaccorsi A, Hermsmeyer K, Aprigliano O, Smith CB, Bohr DF. Mechanism of potassium-induced relaxation in arterial smooth muscle. Blood Vessels. 1977;14:261–276. - PubMed
-
- Hamill OP, Marty A, Neher B, Sigworth FJ. Improved patch-clamp techniques for high resolution current recording from cells and cell-free membrane patches. Pflügers Archiv. 1981;391:85–100. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous