

Beta2-adrenergic receptor overexpression in the developing mouse heart: evidence for targeted modulation of ion channels
- PMID: 10066919
- PMCID: PMC2269209
- DOI: 10.1111/j.1469-7793.1999.019aa.x
Beta2-adrenergic receptor overexpression in the developing mouse heart: evidence for targeted modulation of ion channels
Abstract
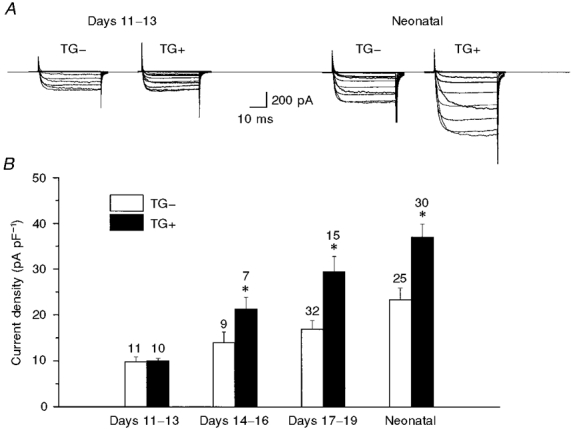
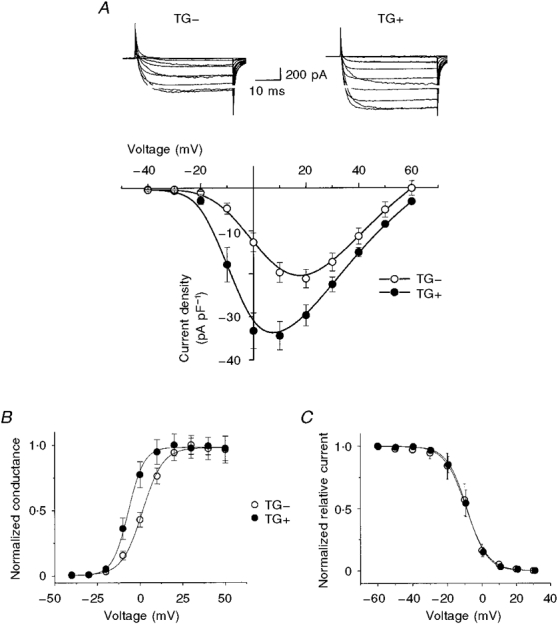
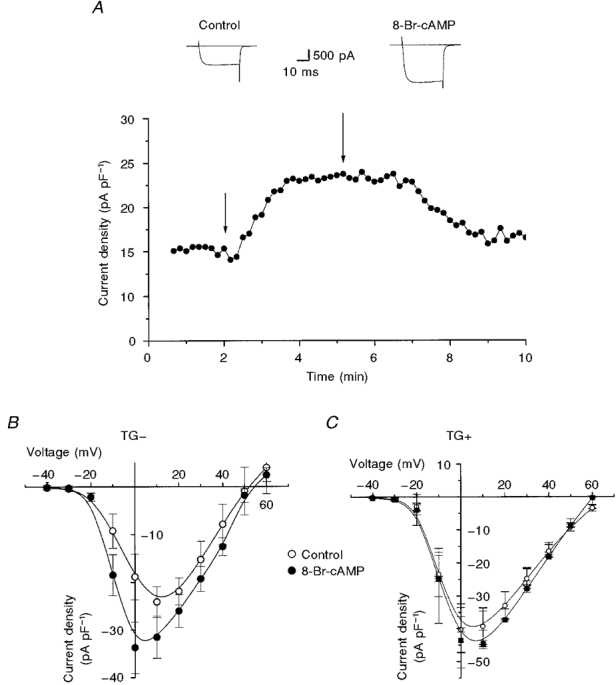
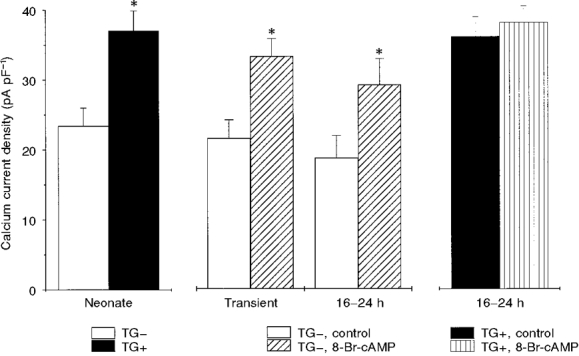
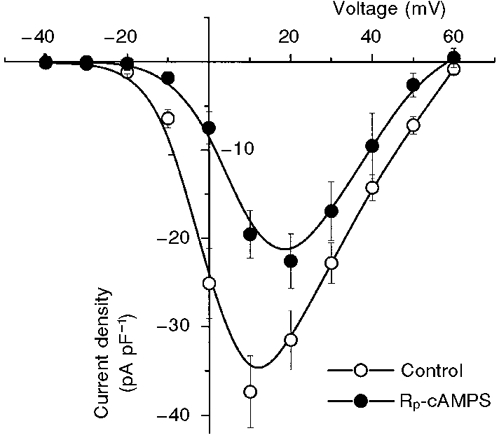
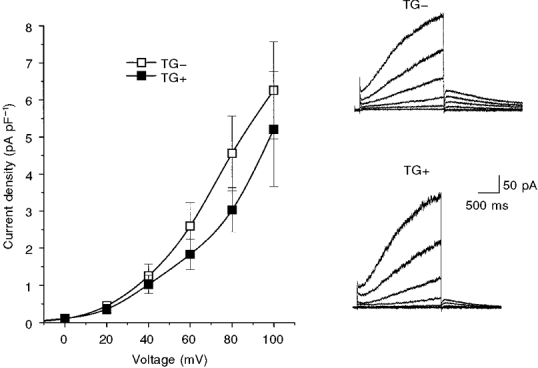
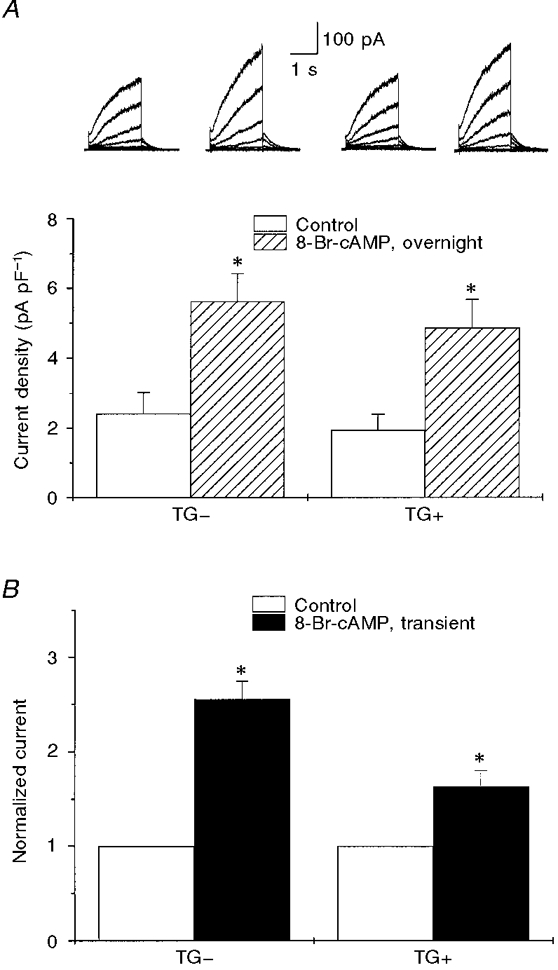
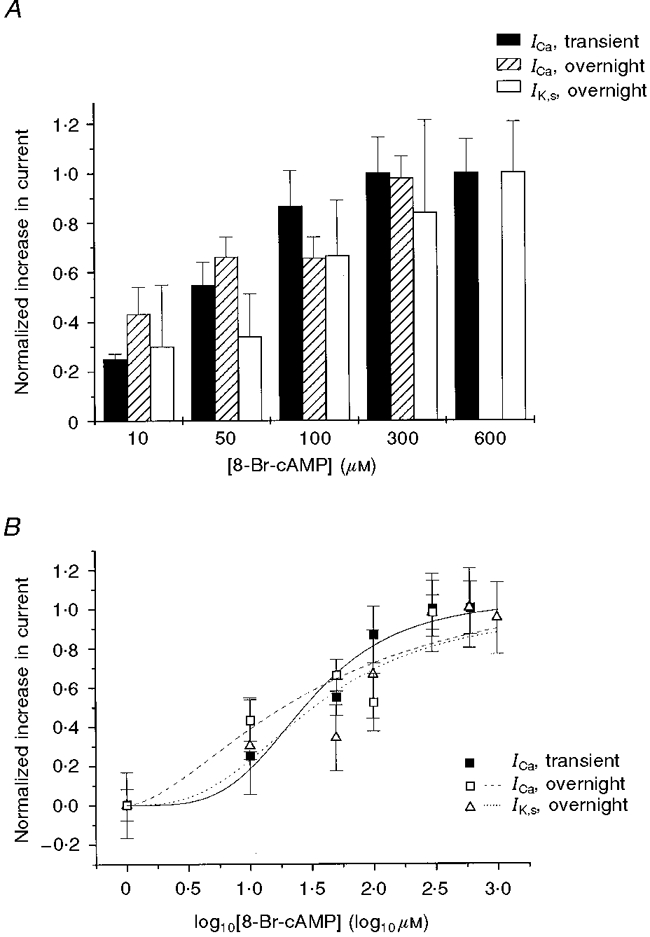
1. We studied the effect of overexpression of the beta2-adrenergic receptor (beta2-AR) in the heart on ion channel currents in single cells isolated from hearts of fetal and neonatal transgenic and wild-type mice. The beta2-AR transgene construct was under the control of the murine alpha-myosin heavy chain (alpha-MHC) promoter, and ion channel activity was measured at distinct developmental stages using whole-cell and perforated patch clamp techniques. 2. We found no change in L-type Ca2+ channel current (ICa) density in early embryonic stages (E11-13) of beta2-AR transgenic positive (TG+) mice, but significant increases in ICa density in intermediate (E14-16, 152 %) and late (E17-19, 173.7 %) fetal and neonatal (1 day post partum, 161 %) TG+ compared with transgenic negative (TG-) mice. This increase in ICa was accompanied by a negative shift in the peak of the current-voltage relationship in TG+ mice. 3. Transient (< 3 min) or prolonged (16-24 h) exposure of TG- neonatal stage myocytes to 8-Br-cAMP (300 microM) increased ICa density and caused a shift in the current-voltage relationship to a similar extent to that seen in TG+ mice. In TG+ myocytes, 8-Br-cAMP had no effect. Exposure of TG+ cells to Rp-cAMPS reversed both the shift in voltage dependence and reduced the peak current density observed in these myocytes. We concluded from these results that the L-type Ca2+ channel is maximally modulated by cAMP-dependent protein kinase (PKA) in TG+ mice and that the alpha-MHC promoter is functional in the ventricle as early as embryonic day 14. 4. In contrast, we found that slow delayed rectifier K+ channels were not changed significantly at any of the developmental stages studied by the overexpression of beta2-ARs compared with TG- mice. The sensitivity of murine slow delayed rectifier K+ channels to cAMP was tested by both transient and prolonged exposure to 8-Br-cAMP (300 microM), which increased the slow delayed rectifier K+ channel current (IK,s) density to a similar extent in both TG- and TG+ neonatal myocytes. In addition, we found that there was no difference in the concentration dependence of the response of ICa and IK,s to 8-Br-cAMP. 5. Thus, overexpression of the beta2-AR in the heart results in distinct modulation of ICa, but not IK,s, and this is not due to differences in the 8-Br-cAMP sensitivity of the two channels. Instead, these results are consistent with both compartmentalization of beta2-AR-controlled cAMP and distinct localization of L-type Ca2+ and slow delayed rectifier K+ channels. This cAMP is targeted preferentially to the L-type Ca2+ channel and is not accessible to the slow delayed rectifier K+ channel.
Figures
References
-
- Aass H, Skomedal T, Osnes JB. Increase of cyclic AMP in subcellular fractions of rat heart muscle after β-adrenergic stimulation: prenalterol and isoprenaline caused different distribution of bound cyclic AMP. Journal of Molecular and Cellular Cardiology. 1988;20:847–860. - PubMed
-
- An RH, Davies MP, Doevendans PA, Kubalak SW, Bangalore R, Chien KR, Kass RS. Developmental changes in β-adrenergic modulation of L-type calcium channels in embryonic mouse heart. Circulation Research. 1996;78:371–378. - PubMed
-
- Bond RA. Do recent operational studies indicate that a single state model is no longer applicable to G protein-coupled receptors? Annals of the New York Academy of Sciences. 1997;812:92–97. - PubMed
-
- Bond RA, Leff P, Johnson TD, Milano CA, Rockman HA, McMinn TR, Apparsundaram S, Hyek MF, Kenakin TP, Allen LF. Physiological effects of inverse agonists in transgenic mice with myocardial overexpression of the β2-adrenoceptor. Nature. 1995;374:272–276. - PubMed
-
- Bristow MR, Ginsburg R, Minobe W, Cubicciotti RS, Sageman WS, Lurie K, Billingham ME, Harrison DC, Stinson EB. Decreased catecholamine sensitivity and β-adrenergic-receptor density in failing human hearts. New England Journal of Medicine. 1982;307:205–211. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous