

GCS1, an Arf guanosine triphosphatase-activating protein in Saccharomyces cerevisiae, is required for normal actin cytoskeletal organization in vivo and stimulates actin polymerization in vitro
- PMID: 10069805
- PMCID: PMC25189
- DOI: 10.1091/mbc.10.3.581
GCS1, an Arf guanosine triphosphatase-activating protein in Saccharomyces cerevisiae, is required for normal actin cytoskeletal organization in vivo and stimulates actin polymerization in vitro
Abstract
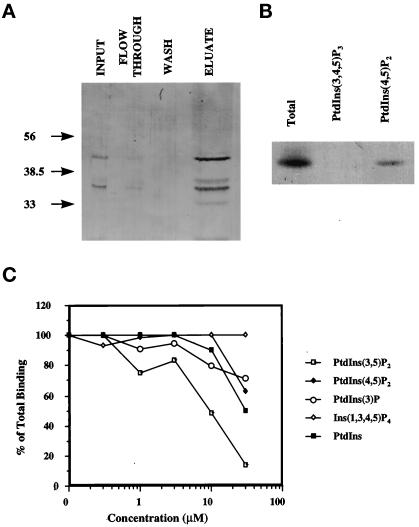
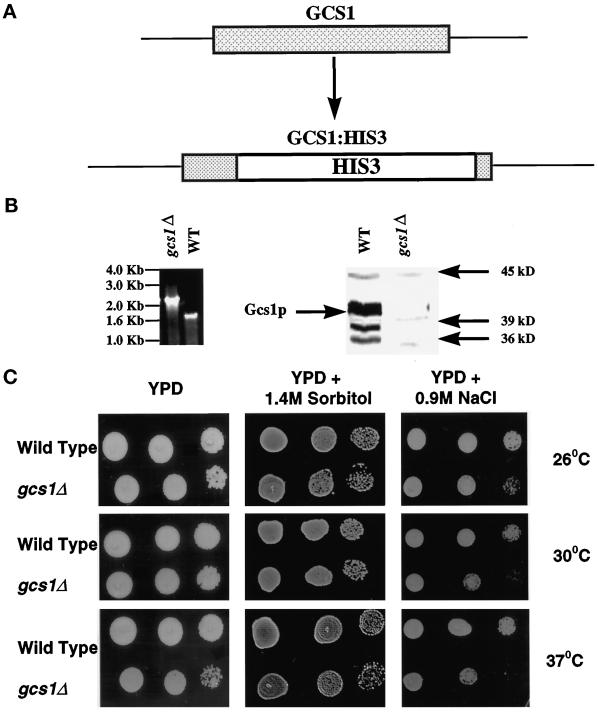
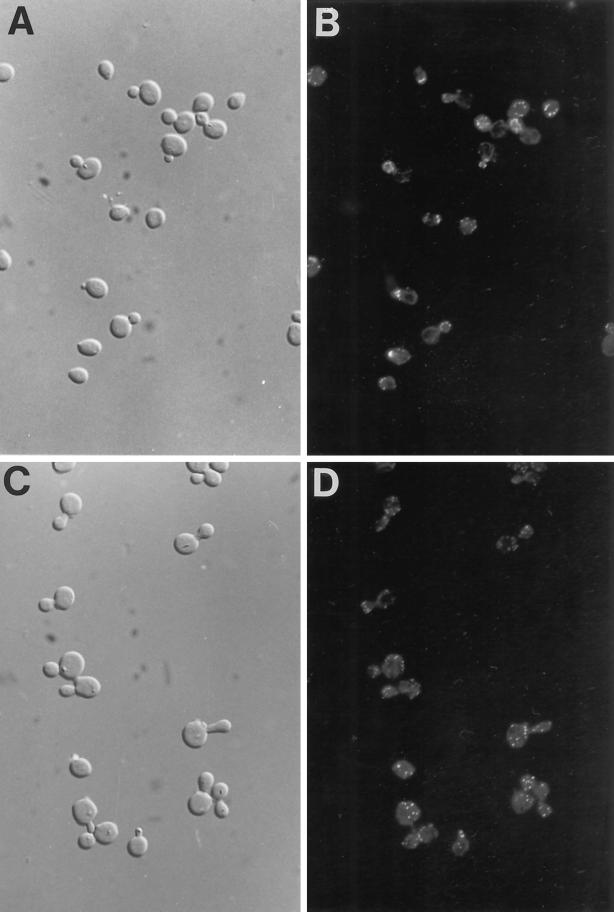
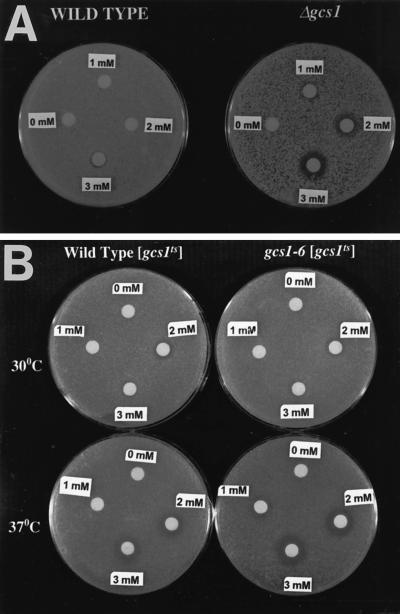
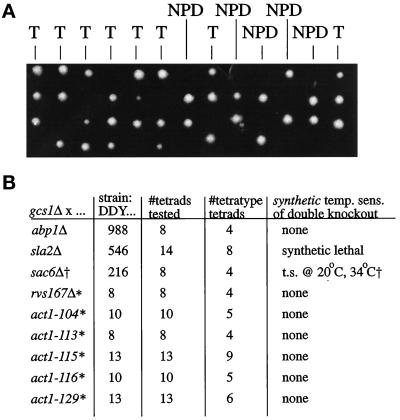
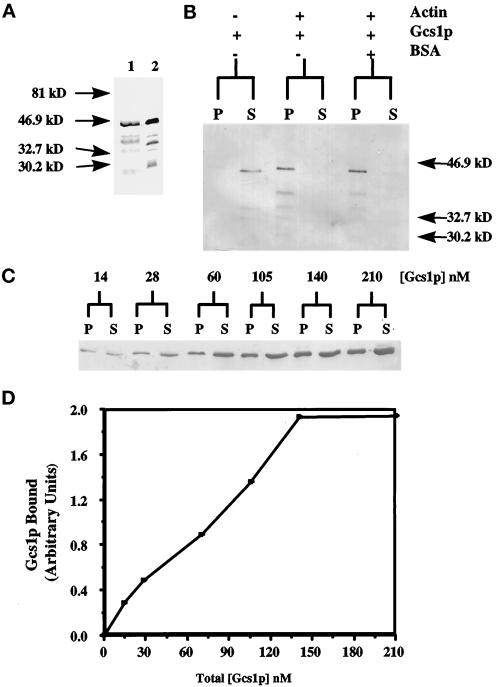
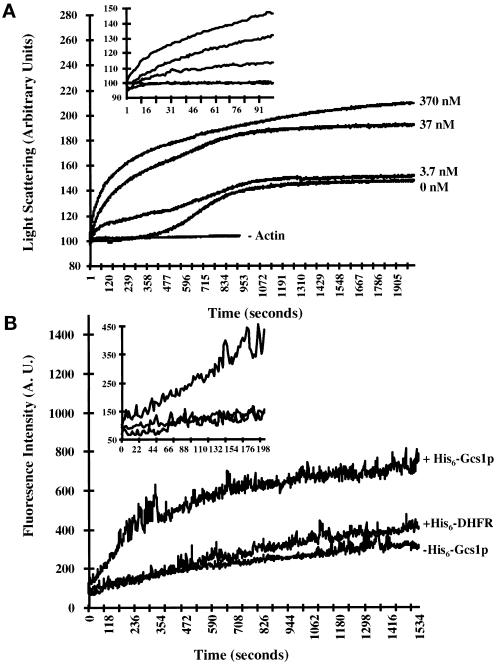
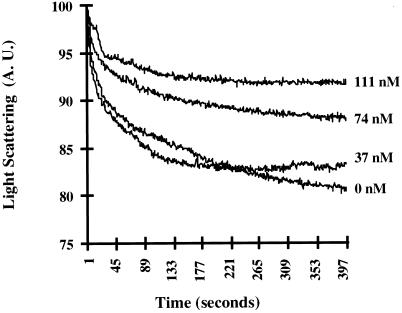
Recent cloning of a rat brain phosphatidylinositol 3,4, 5-trisphosphate binding protein, centaurin alpha, identified a novel gene family based on homology to an amino-terminal zinc-binding domain. In Saccharomyces cerevisiae, the protein with the highest homology to centaurin alpha is Gcs1p, the product of the GCS1 gene. GCS1 was originally identified as a gene conditionally required for the reentry of cells into the cell cycle after stationary phase growth. Gcs1p was previously characterized as a guanosine triphosphatase-activating protein for the small guanosine triphosphatase Arf1, and gcs1 mutants displayed vesicle-trafficking defects. Here, we have shown that similar to centaurin alpha, recombinant Gcs1p bound phosphoinositide-based affinity resins with high affinity and specificity. A novel GCS1 disruption strain (gcs1Delta) exhibited morphological defects, as well as mislocalization of cortical actin patches. gcs1Delta was hypersensitive to the actin monomer-sequestering drug, latrunculin-B. Synthetic lethality was observed between null alleles of GCS1 and SLA2, the gene encoding a protein involved in stabilization of the actin cytoskeleton. In addition, synthetic growth defects were observed between null alleles of GCS1 and SAC6, the gene encoding the yeast fimbrin homologue. Recombinant Gcs1p bound to actin filaments, stimulated actin polymerization, and inhibited actin depolymerization in vitro. These data provide in vivo and in vitro evidence that Gcs1p interacts directly with the actin cytoskeleton in S. cerevisiae.
Figures

References
-
- Adams AE, Botstein D, Drubin DG. Requirement of yeast fimbrin for actin organization and morphogenesis in vivo. Nature. 1991;354:404–408. - PubMed
-
- Antonny B, Huber I, Paris S, Chabre M, Cassel D. Activation of ADP-ribosylation factor 1 GTPase-activating protein by phosphatidylcholine-derived diacylglycerols. J Biol Chem. 1997;272:30848–30851. - PubMed
-
- Ayscough KR, Drubin DG. ACTIN: general principles from studies in yeast. Annu Rev Cell Dev Biol. 1996;12:129–160. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
