

Binding of hnRNP H to an exonic splicing silencer is involved in the regulation of alternative splicing of the rat beta-tropomyosin gene
- PMID: 10072387
- PMCID: PMC316507
- DOI: 10.1101/gad.13.5.593
Binding of hnRNP H to an exonic splicing silencer is involved in the regulation of alternative splicing of the rat beta-tropomyosin gene
Abstract
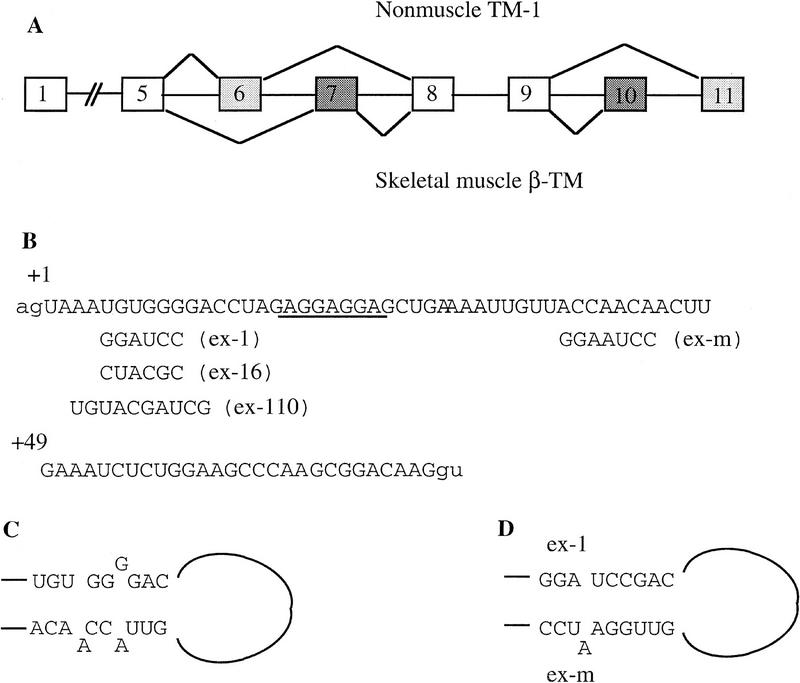
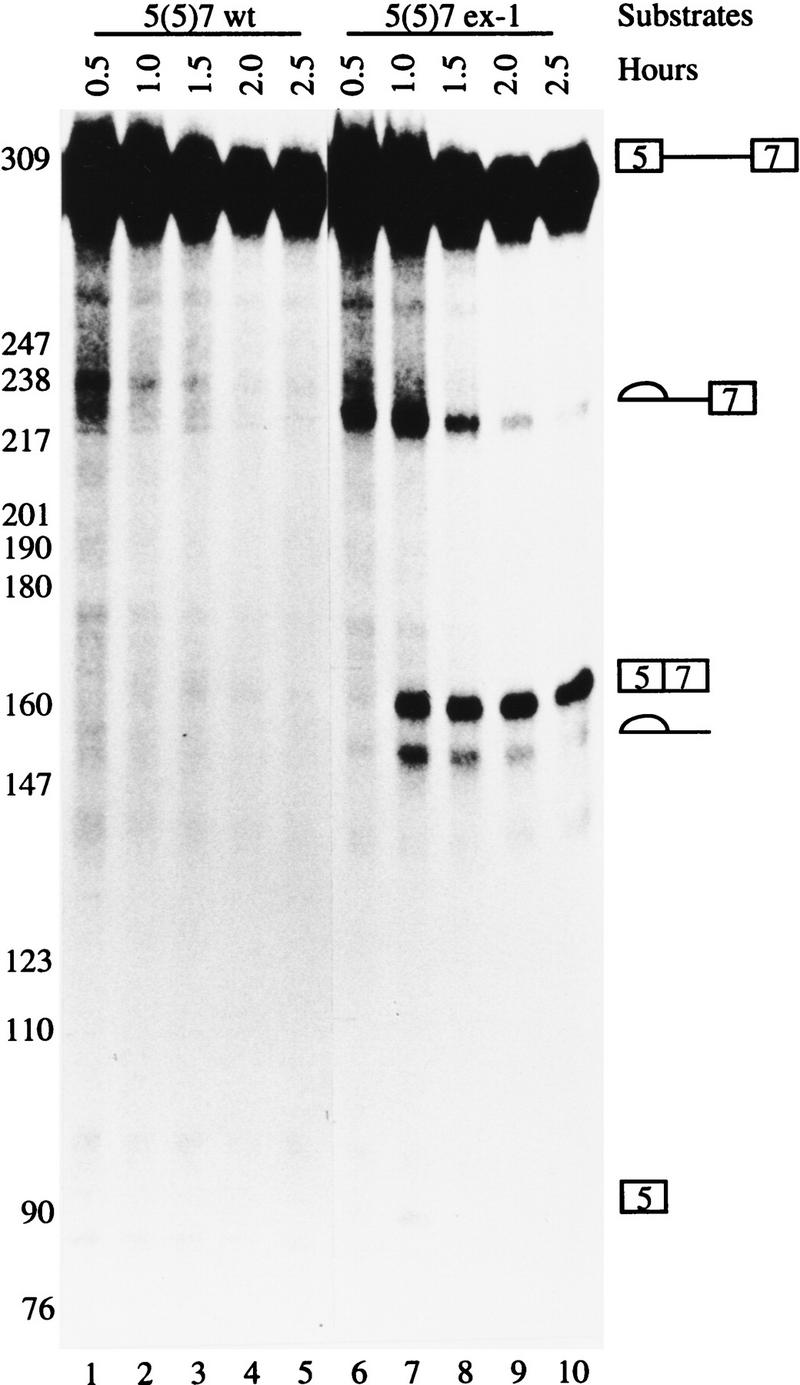
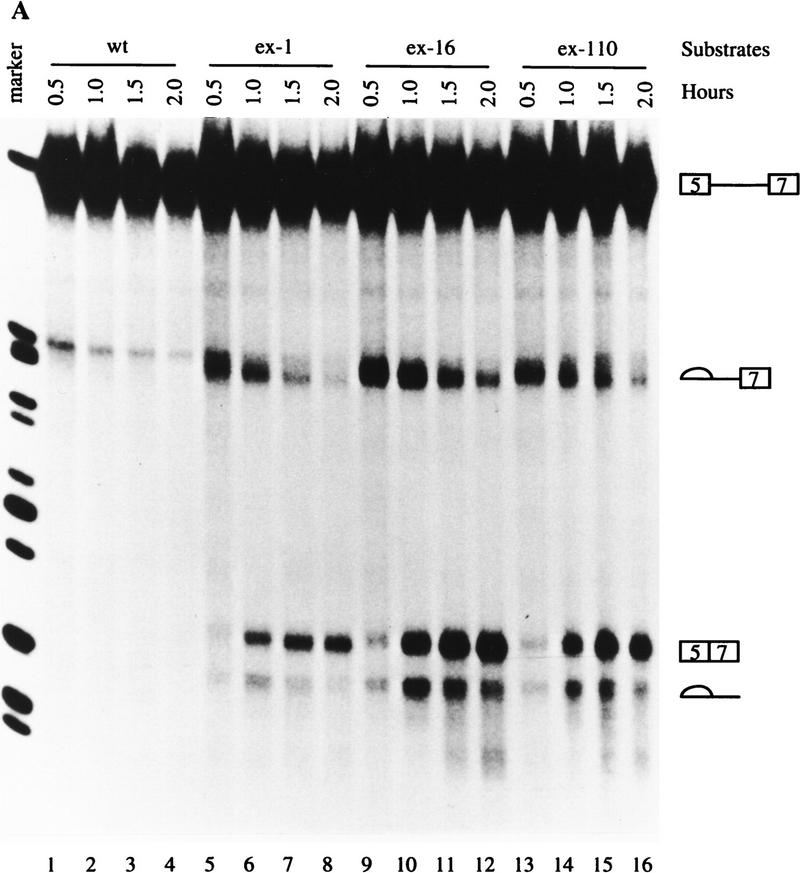
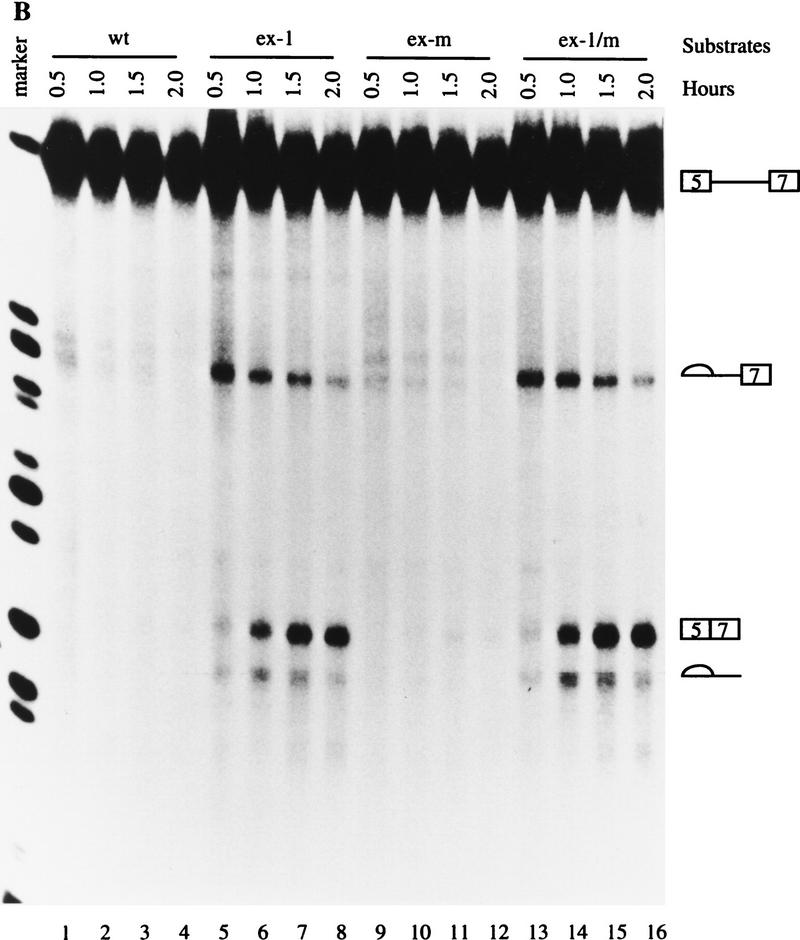
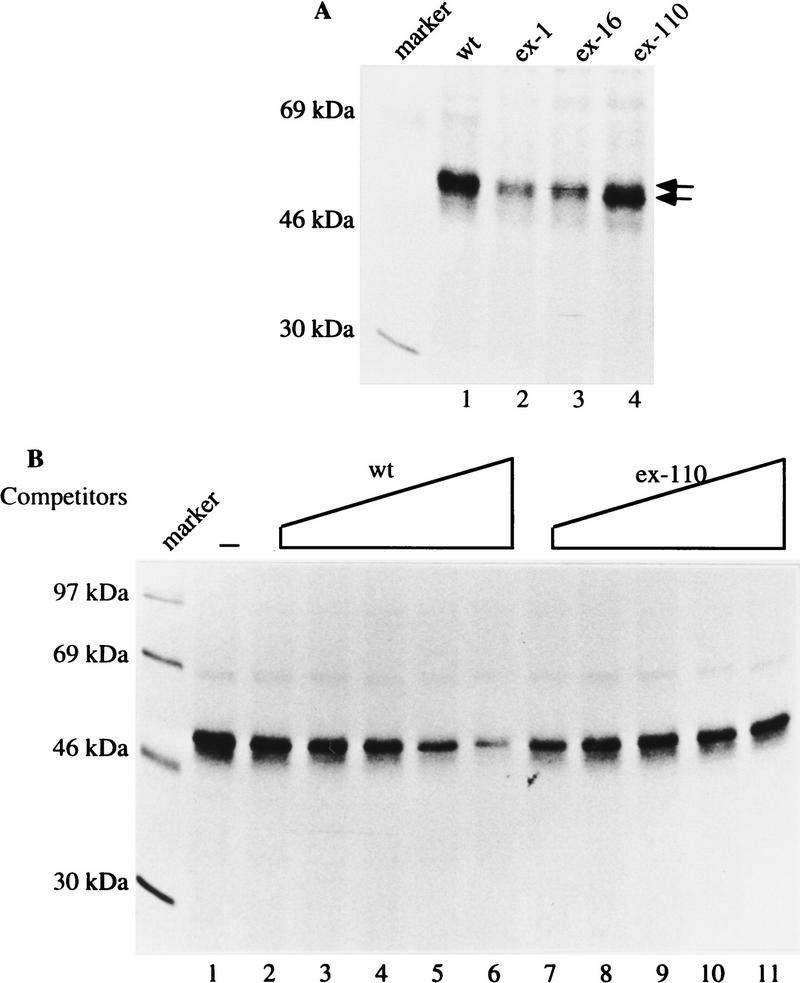
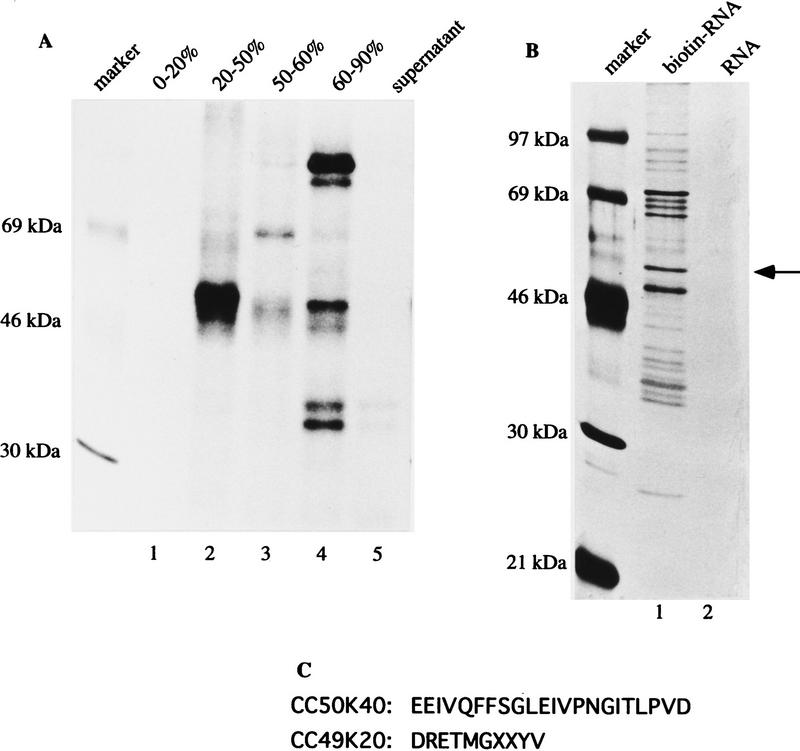
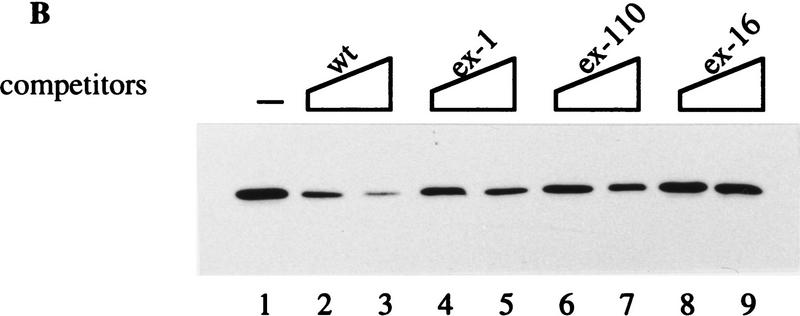
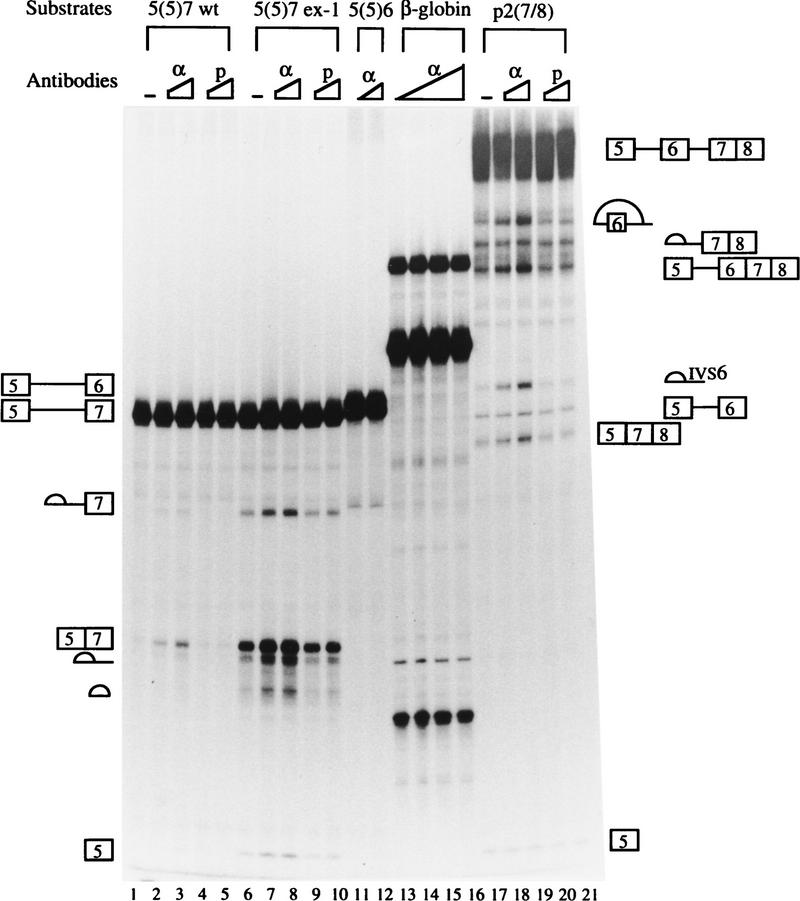
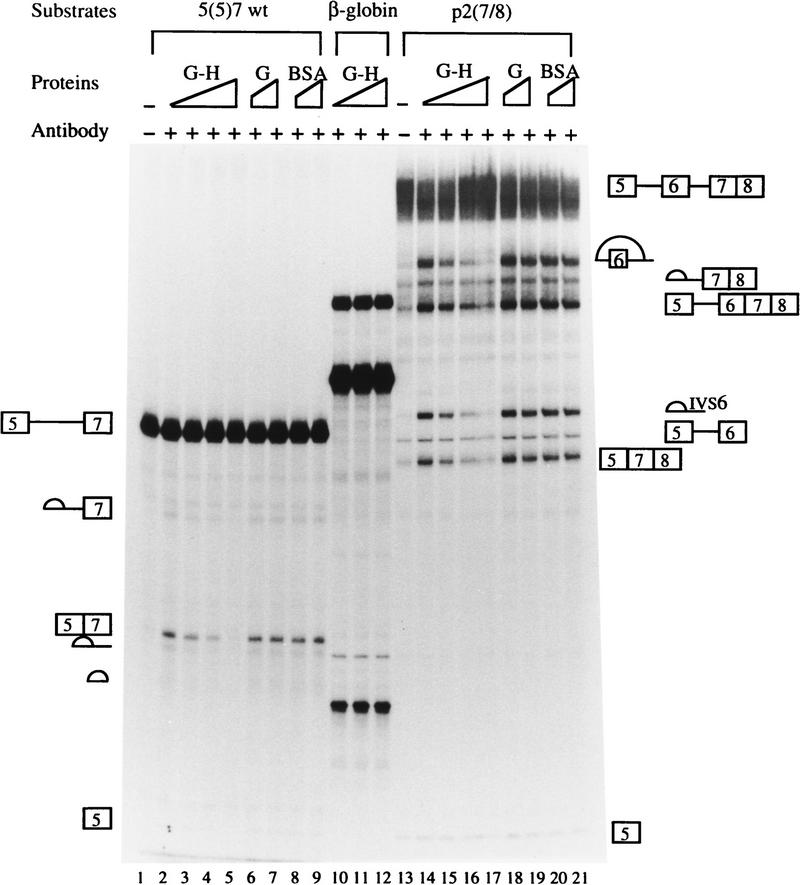
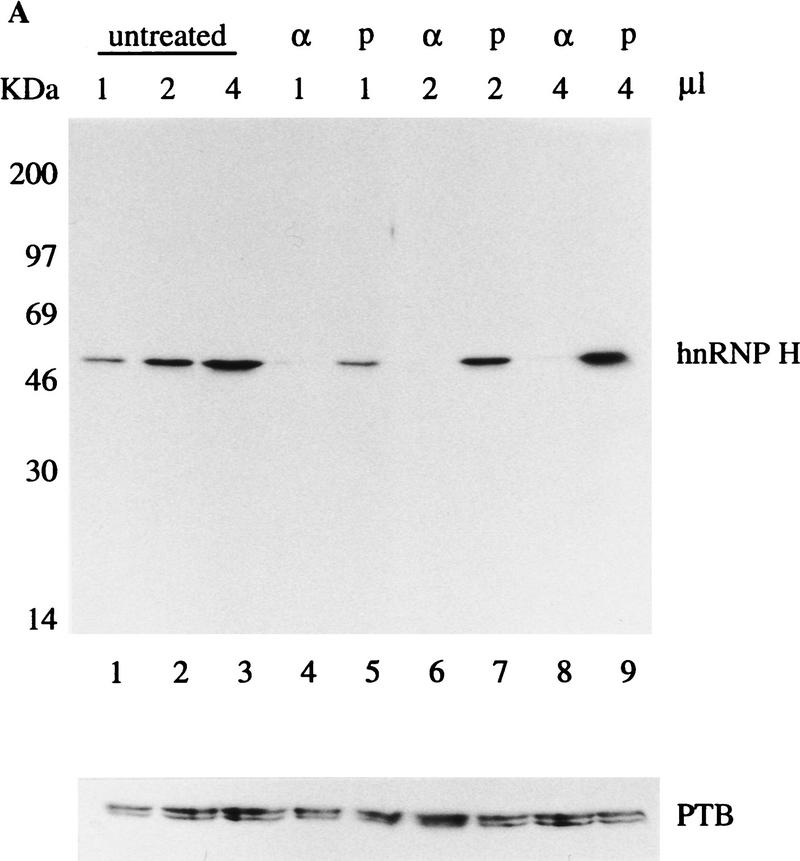
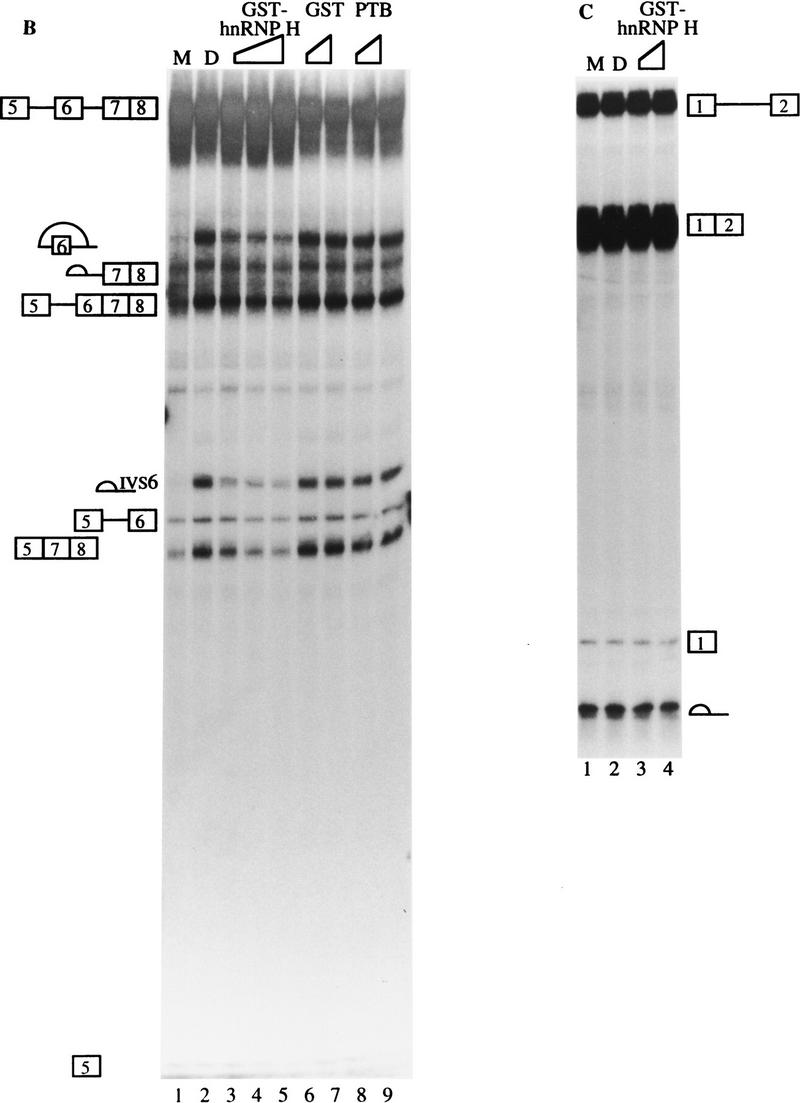
In the rat beta-tropomyosin (beta-TM) gene, exons 6 and 7 are spliced alternatively in a mutually exclusive manner. Exon 6 is included in mRNA encoding nonmuscle TM-1, whereas exon 7 is used in mRNA encoding skeletal muscle beta-TM. Previously, we demonstrated that a six nucleotide mutation at the 5' end of exon 7, designated as ex-1, activated exon 7 splicing in nonmuscle cells. In this study, we show that the activating effect of this mutation is not the result of creating an exonic splicing enhancer (ESE) or disrupting a putative secondary structure. The sequence in exon 7 acts as a bona fide exonic splicing silencer (ESS), which is bound specifically by a trans-acting factor. Isolation and peptide sequencing reveal that this factor is hnRNP H, a member of the heterogeneous nuclear ribonucleoprotein (hnRNP) family. Binding of hnRNP H correlates with the ESS activity. Furthermore, addition of antibodies that specifically recognizes hnRNP H to the splicing reactions or partial depletion of hnRNP H from nuclear extract activates exon 7 splicing in vitro and this effect can be reversed by addition of purified recombinant hnRNP H. These results indicate that hnRNP H participates in exclusion of exon 7 in nonmuscle cells. The involvement of hnRNP H in the activity of an ESS may represent a prototype for the regulation of tissue- and developmental-specific alternative splicing.
Figures


References
-
- Adams MD, Rudner DZ, Rio DC. Biochemistry and regulation of pre-mRNA splicing. Curr Opin Cell Biol. 1996;8:331–339. - PubMed
-
- Black D. Activation of c-src neuron-specific splicing by an unusual RNA element in vitro and in vivo. Cell. 1992;69:795–807. - PubMed
-
- Boggs RT, Gregor P, Idriss S, Belote JM, McKeown M. Regulation of sexual differentiation in D. melanogaster via alternative splicing of RNA from the transformer gene. Cell. 1987;50:739–747. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources