

Tyrosine aminotransferase catalyzes the final step of methionine recycling in Klebsiella pneumoniae
- PMID: 10074065
- PMCID: PMC93571
- DOI: 10.1128/JB.181.6.1739-1747.1999
Tyrosine aminotransferase catalyzes the final step of methionine recycling in Klebsiella pneumoniae
Abstract
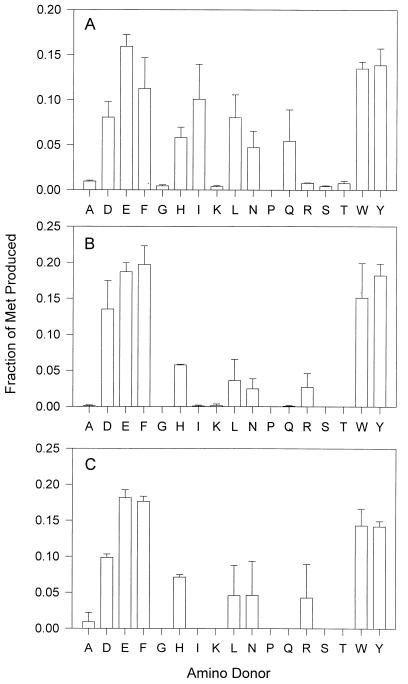
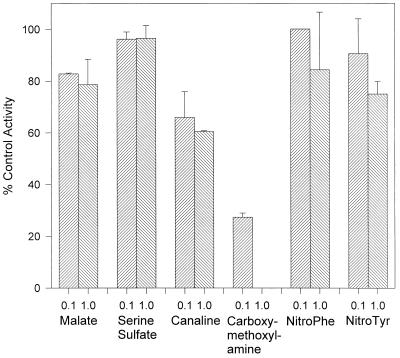
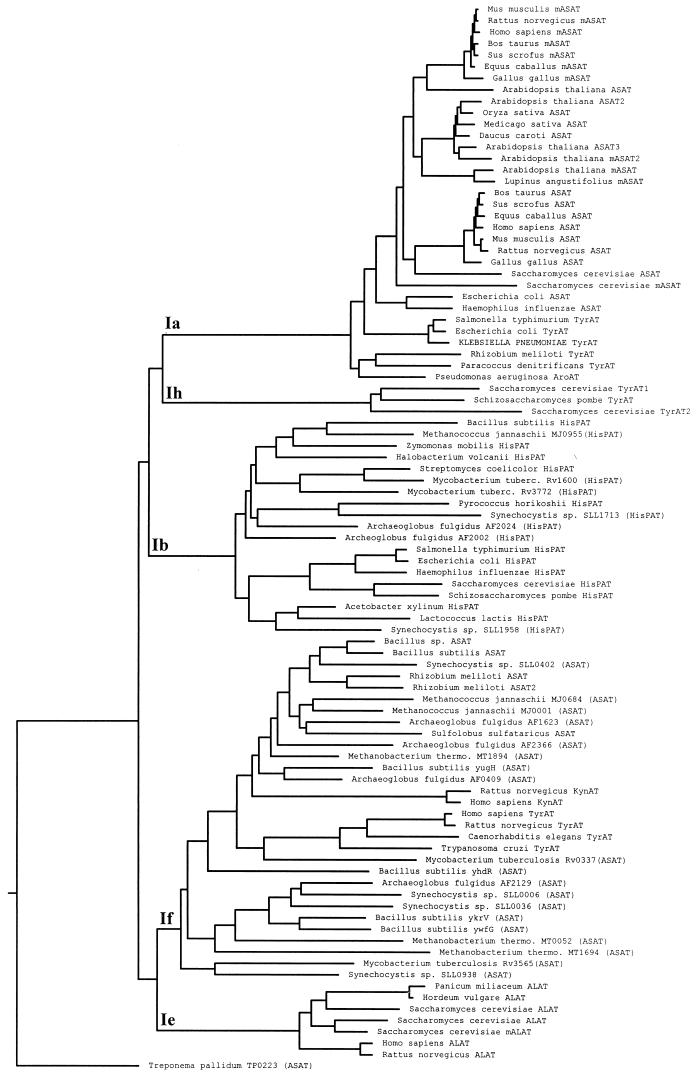
An aminotransferase which catalyzes the final step in methionine recycling from methylthioadenosine, the conversion of alpha-ketomethiobutyrate to methionine, has been purified from Klebsiella pneumoniae and characterized. The enzyme was found to be a homodimer of 45-kDa subunits, and it catalyzed methionine formation primarily using aromatic amino acids and glutamate as the amino donors. Histidine, leucine, asparagine, and arginine were also functional amino donors but to a lesser extent. The N-terminal amino acid sequence of the enzyme was determined and found to be almost identical to the N-terminal sequence of both the Escherichia coli and Salmonella typhimurium tyrosine aminotransferases (tyrB gene products). The structural gene for the tyrosine aminotransferase was cloned from K. pneumoniae and expressed in E. coli. The deduced amino acid sequence displayed 83, 80, 38, and 34% identity to the tyrosine aminotransferases from E. coli, S. typhimurium, Paracoccus denitrificans, and Rhizobium meliloti, respectively, but it showed less than 13% identity to any characterized eukaryotic tyrosine aminotransferase. Structural motifs around key invariant residues placed the K. pneumoniae enzyme within the Ia subfamily of aminotransferases. Kinetic analysis of the aminotransferase showed that reactions of an aromatic amino acid with alpha-ketomethiobutyrate and of glutamate with alpha-ketomethiobutyrate proceed as favorably as the well-known reactions of tyrosine with alpha-ketoglutarate and tyrosine with oxaloacetate normally associated with tyrosine aminotransferases. The aminotransferase was inhibited by the aminooxy compounds canaline and carboxymethoxylamine but not by substrate analogues, such as nitrotyrosine or nitrophenylalanine.
Figures


References
-
- Backlund P S, Chang C P, Smith R A. Identification of 2-keto-4-methylthiobutyrate as an intermediate compound in methionine synthesis from 5′-methylthioadenosine. J Biol Chem. 1982;257:4196–4202. - PubMed
-
- Backlund P S, Smith R A. Methionine synthesis from 5′-methylthioadenosine in rat liver. J Biol Chem. 1980;256:1533–1535. - PubMed
-
- Berger B J, Dai W W, Wilson J. Methionine formation from α-ketomethiobutyrate in the trypanosomatid Crithidia fasciculata. FEMS Microbiol Lett. 1998;165:305–312. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
