

Purification and characterization of two extremely thermostable enzymes, phosphate acetyltransferase and acetate kinase, from the hyperthermophilic eubacterium Thermotoga maritima
- PMID: 10074080
- PMCID: PMC93586
- DOI: 10.1128/JB.181.6.1861-1867.1999
Purification and characterization of two extremely thermostable enzymes, phosphate acetyltransferase and acetate kinase, from the hyperthermophilic eubacterium Thermotoga maritima
Abstract
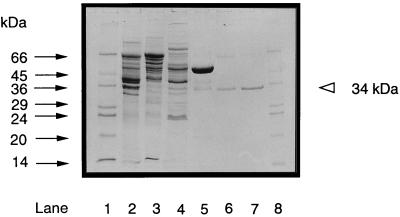
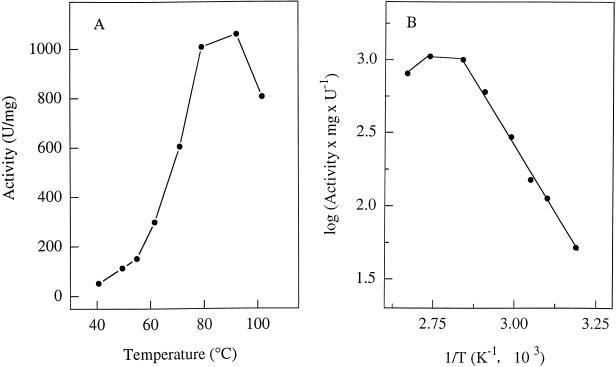
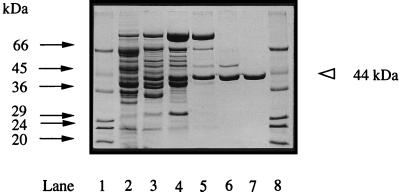
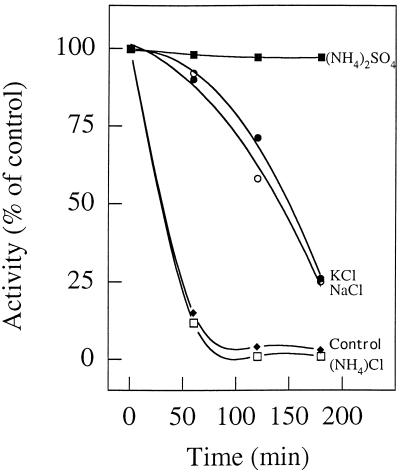
Phosphate acetyltransferase (PTA) and acetate kinase (AK) of the hyperthermophilic eubacterium Thermotoga maritima have been purified 1,500- and 250-fold, respectively, to apparent homogeneity. PTA had an apparent molecular mass of 170 kDa and was composed of one subunit with a molecular mass of 34 kDa, suggesting a homotetramer (alpha4) structure. The N-terminal amino acid sequence showed significant identity to that of phosphate butyryltransferases from Clostridium acetobutylicum rather than to those of known phosphate acetyltransferases. The kinetic constants of the reversible enzyme reaction (acetyl-CoA + Pi -->/<-- acetyl phosphate + CoA) were determined at the pH optimum of pH 6.5. The apparent Km values for acetyl-CoA, Pi, acetyl phosphate, and coenzyme A (CoA) were 23, 110, 24, and 30 microM, respectively; the apparent Vmax values (at 55 degrees C) were 260 U/mg (acetyl phosphate formation) and 570 U/mg (acetyl-CoA formation). In addition to acetyl-CoA (100%), the enzyme accepted propionyl-CoA (60%) and butyryl-CoA (30%). The enzyme had a temperature optimum at 90 degrees C and was not inactivated by heat upon incubation at 80 degrees C for more than 2 h. AK had an apparent molecular mass of 90 kDa and consisted of one 44-kDa subunit, indicating a homodimer (alpha2) structure. The N-terminal amino acid sequence showed significant similarity to those of all known acetate kinases from eubacteria as well that of the archaeon Methanosarcina thermophila. The kinetic constants of the reversible enzyme reaction (acetyl phosphate + ADP -->/<-- acetate + ATP) were determined at the pH optimum of pH 7.0. The apparent Km values for acetyl phosphate, ADP, acetate, and ATP were 0.44, 3, 40, and 0.7 mM, respectively; the apparent Vmax values (at 50 degrees C) were 2,600 U/mg (acetate formation) and 1,800 U/mg (acetyl phosphate formation). AK phosphorylated propionate (54%) in addition to acetate (100%) and used GTP (100%), ITP (163%), UTP (56%), and CTP (21%) as phosphoryl donors in addition to ATP (100%). Divalent cations were required for activity, with Mn2+ and Mg2+ being most effective. The enzyme had a temperature optimum at 90 degrees C and was stabilized against heat inactivation by salts. In the presence of (NH4)2SO4 (1 M), which was most effective, the enzyme did not lose activity upon incubation at 100 degrees C for 3 h. The temperature optimum at 90 degrees C and the high thermostability of both PTA and AK are in accordance with their physiological function under hyperthermophilic conditions.
Figures
References
-
- Aceti D J, Ferry J G. Purification and characterization of acetate kinase from acetate-grown Methanosarcina thermophila. Evidence for regulation of synthesis. J Biol Chem. 1988;263:15444–15448. - PubMed
-
- Adams M W W, editor. Enzymes and proteins from hyperthermophilic microorganisms. Adv Protein Chem. 1996;48:1–509.
-
- Bergmeyer H U, Holz G, Klotzsch H, Lang G. Phosphotransacetylase aus Clostridium kluyveri. Züchtung des Bacteriums, Isolierung, Kristallisation und Eigenschaften des Enzymes. Biochem Z. 1963;338:114–121. - PubMed
-
- Bowmann C M, Valdez R O, Nishimura J S. Acetate kinase from Veillonella alcalescens. Regulations of enzyme activity by succinate and substrates. J Biol Chem. 1976;251:3117–3121. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
