

Mutagenic analysis of the 3' cis-acting elements of the rubella virus genome
- PMID: 10074193
- PMCID: PMC104103
- DOI: 10.1128/JVI.73.4.3386-3403.1999
Mutagenic analysis of the 3' cis-acting elements of the rubella virus genome
Abstract
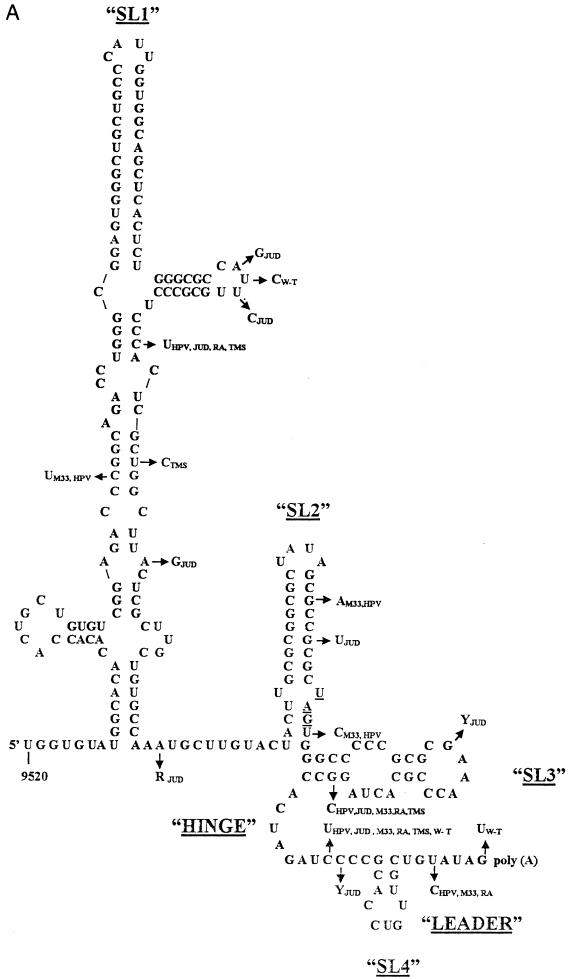
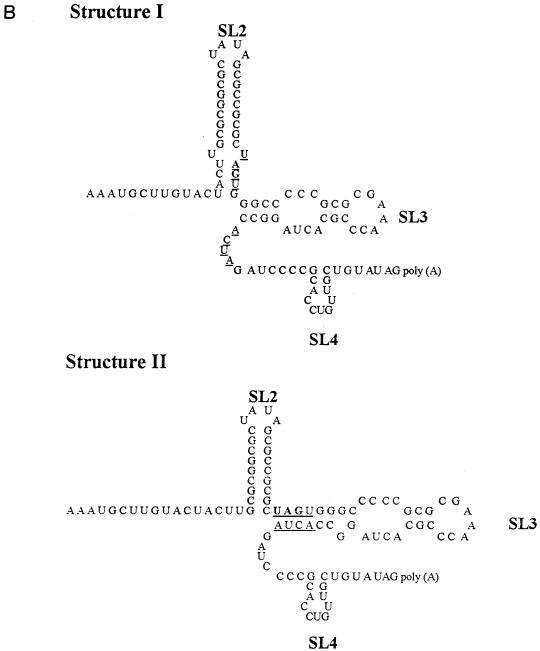
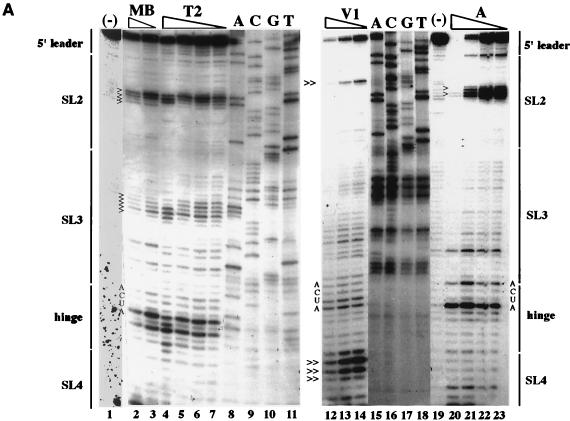
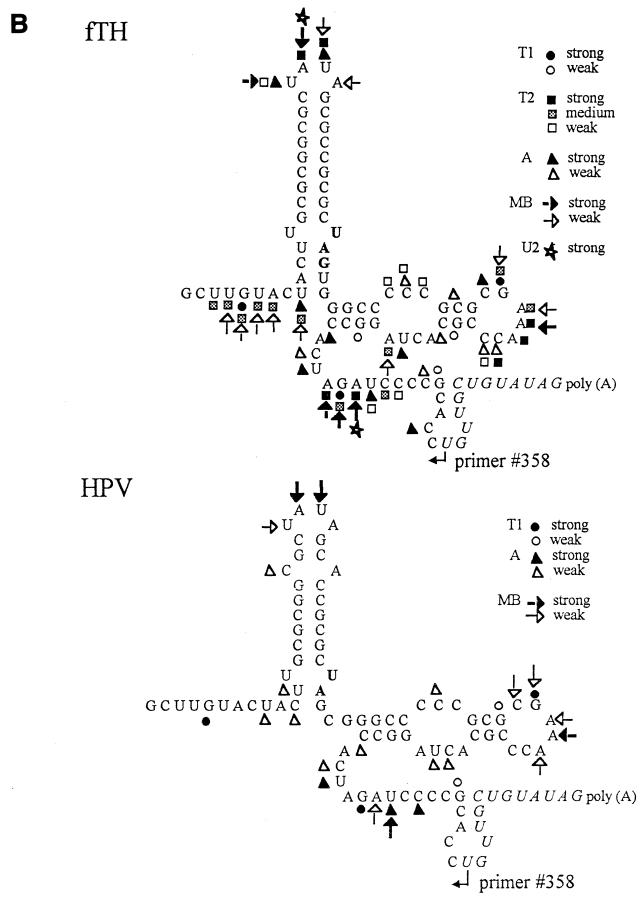
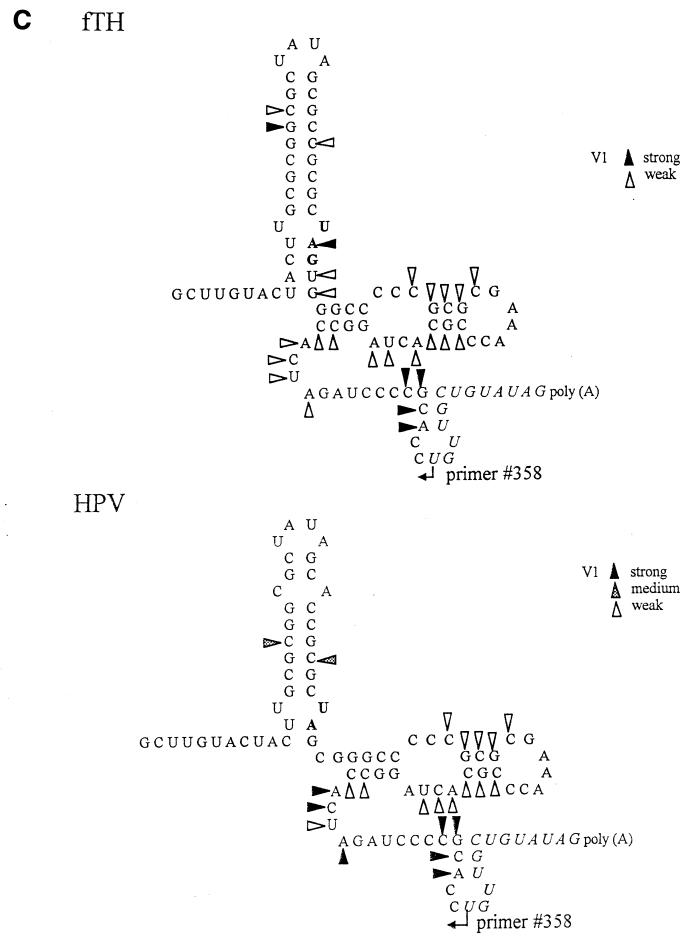
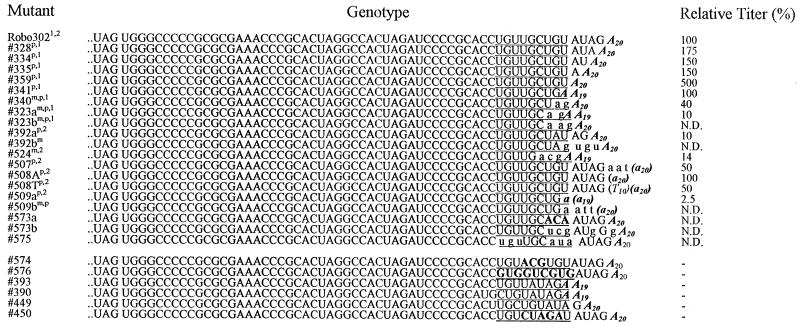
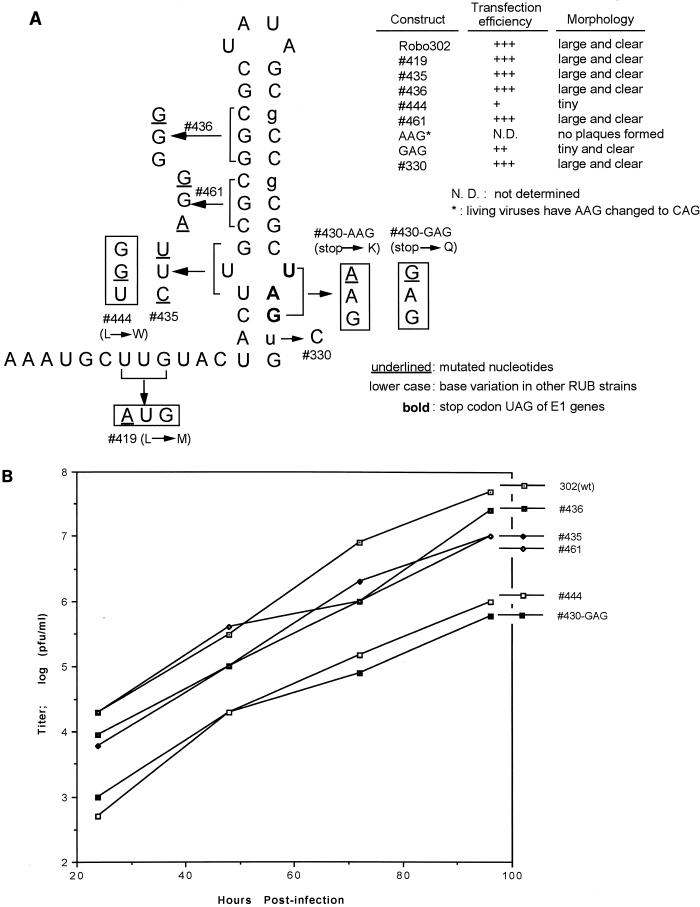
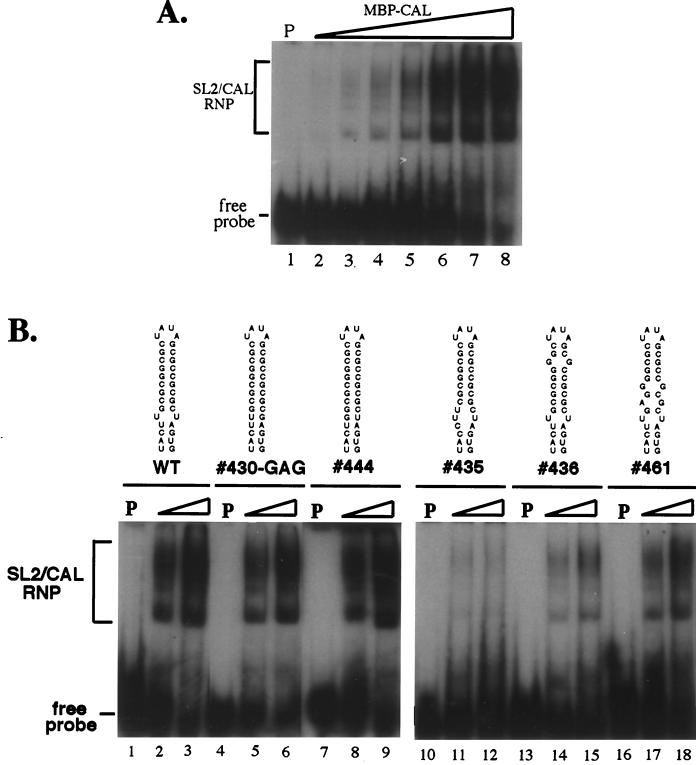
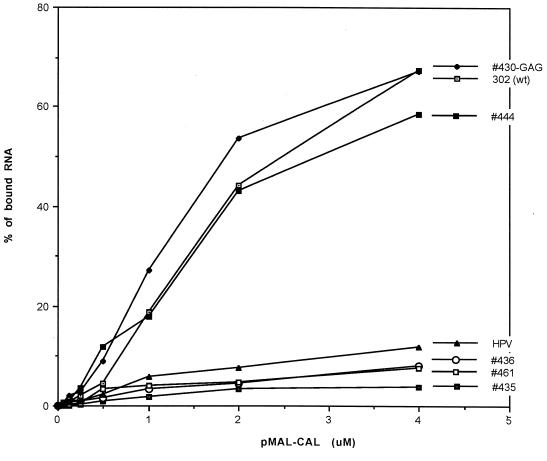
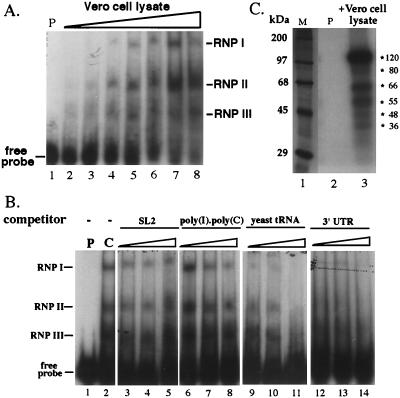
Thermodynamically predicted secondary structure analysis of the 3'-terminal 305 nucleotides (nt) of the rubella virus (RUB) genome, a region conserved in all RUB defective interfering RNAs, revealed four stem-loop (SL) structures; SL1 and SL2 are both located in the E1 coding region, while SL3 and SL4 are within the 59-nt 3' untranslated region (UTR) preceding the poly(A) tract. SL2 is a structure shown to interact with human calreticulin (CAL), an autoantigen potentially involved in RUB RNA replication and pathogenesis. RNase mapping indicated that SL2 and SL3 are in equilibrium between two conformations, in the second of which the previously proposed CAL binding site in SL2, a U-U bulge, is not formed. Site-directed mutagenesis of the 3' UTR with a RUB infectious clone, Robo302, revealed that most of the 3' UTR is required for viral viability except for the 3'-terminal 5 nt and the poly(A) tract, although poly(A) was rapidly regenerated during subsequent replication. Maintenance of the overall SL3 structure, the 11-nt single-stranded sequence between SL3 and SL4, and the sequences forming SL4 were all important for viral viability. Studies on the interaction between host factors and the 3' UTR showed the formation of three RNA-protein complexes by gel mobility shift assay, and UV-induced cross-linking detected six host protein species, with molecular masses of 120, 80, 66, 55, 48, and 36 kDa, interacting with the 3' UTR. Site-directed mutagenesis of SL2 by nucleotide substitutions showed that maintenance of SL2 stem rather than the U-U bulge was critical in CAL binding since mutants having the U-U bulge base paired had a similar binding activity for CAL as the native structure whereas mutants having the SL2 stem destabilized had much lower binding activity. However, all of these mutations gave rise to viable viruses when introduced into Robo302, indicating that binding of CAL to SL2 is independent of viral viability.
Figures


References
-
- Atreya, C. D. Personal communication.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
