

Infectious pancreatic necrosis virus: identification of a VP3-containing ribonucleoprotein core structure and evidence for O-linked glycosylation of the capsid protein VP2
- PMID: 10074207
- PMCID: PMC104117
- DOI: 10.1128/JVI.73.4.3484-3490.1999
Infectious pancreatic necrosis virus: identification of a VP3-containing ribonucleoprotein core structure and evidence for O-linked glycosylation of the capsid protein VP2
Abstract
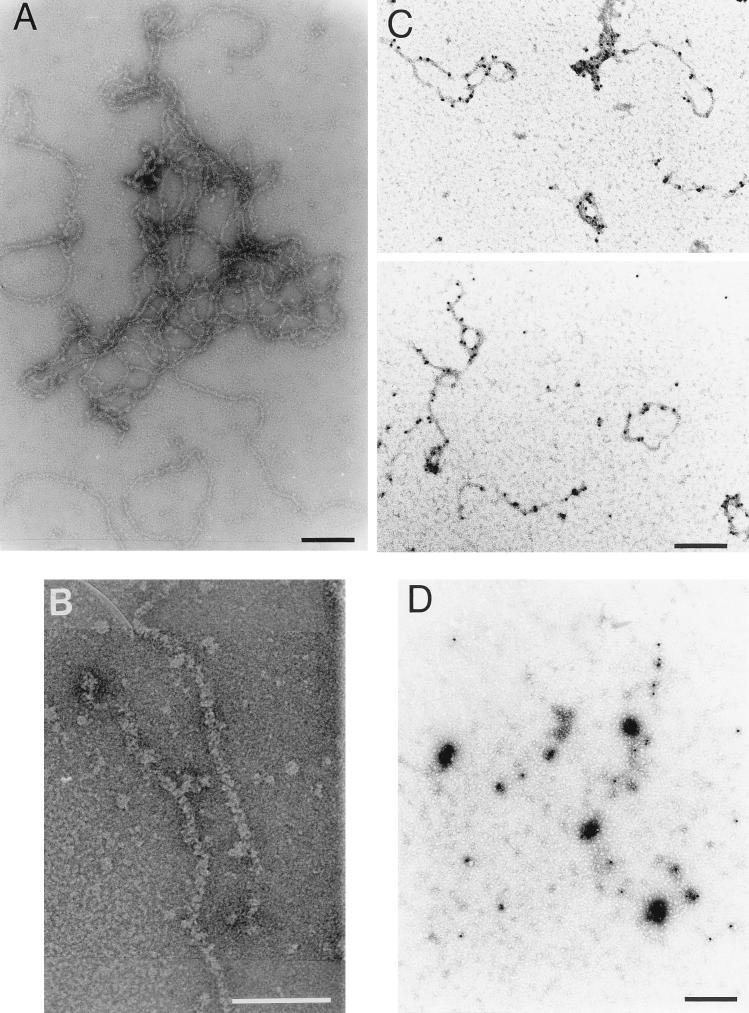
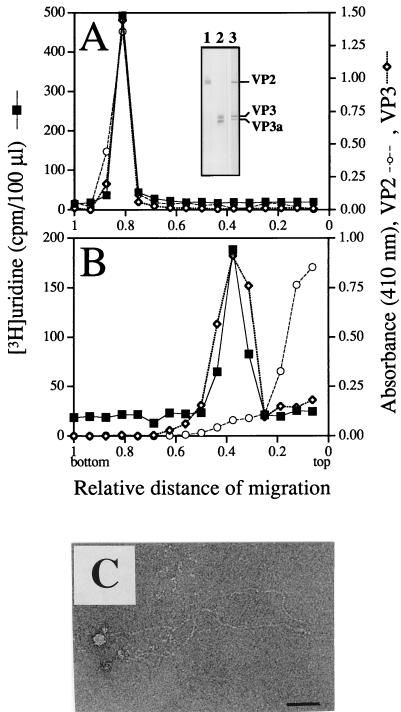
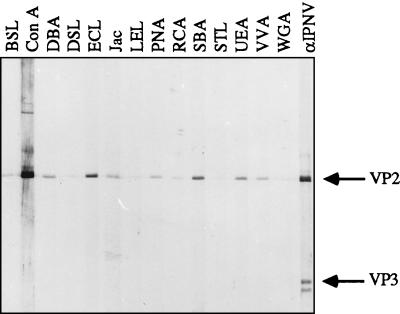
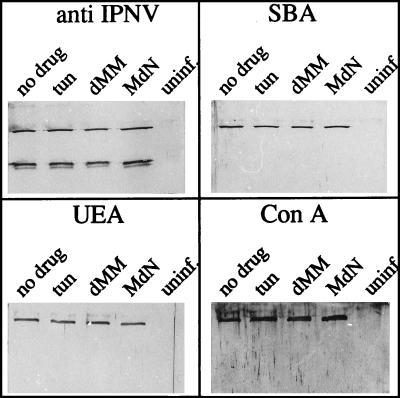
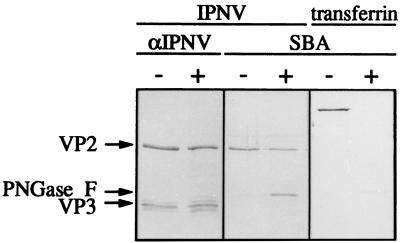
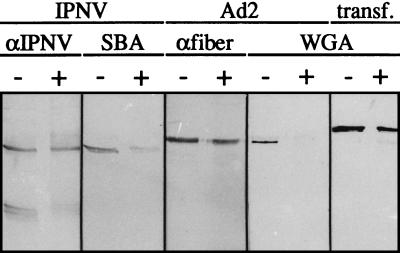
Virions of infectious pancreatic necrosis virus (IPNV) were completely disintegrated upon dialysis against salt-free buffers. Direct visualization of such preparations by electron microscopy revealed 5.0- to 6.5-nm-thick entangled filaments. By using a specific colloidal gold immunolabeling technique, these structures were shown to contain the viral protein VP3. Isolation by sucrose gradient centrifugation of the filaments, followed by serological analysis, demonstrated that the entire VP3 content of the virion was recovered together with the radiolabeled genomic material forming the unique threadlike ribonucleoprotein complexes. In a sensitive blotting assay, the outer capsid component of IPNV, i.e., the major structural protein VP2, was shown to specifically bind lectins recognizing sugar moieties of N-acetylgalactosamine, mannose, and fucose. Three established metabolic inhibitors of N-linked glycosylation did not prevent addition of sugar residues to virions, and enzymatic deglycosylation of isolated virions using N-glycosidase failed to remove sugar residues of VP2 recognized by lectins. However, gentle alkaline beta elimination clearly reduced the ability of lectins to recognize VP2. These results suggest that the glycosylation of VP2 is of the O-linked type when IPNV is propagated in RTG-2 cells.
Figures
References
-
- Bartles J R, Hubbard A L. 125I-wheat germ agglutinin blotting: increased sensitivity with polyvinylpyrrolidone quenching and periodate oxidation/reductive phenylamination. Anal Biochem. 1984;140:284–292. - PubMed
-
- Carlsson A, Kuznar J, Varga M, Everitt E. Purification of infectious pancreatic necrosis virus by anion exchange chromatography increases the specific infectivity. J Virol Methods. 1994;47:27–36. - PubMed
-
- Caswell-Reno P, Reno P W, Nicholson B L. Monoclonal antibodies to infectious pancreatic necrosis virus: analysis of viral epitopes and comparison of different isolates. J Gen Virol. 1986;67:2193–2206. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
