

The cyclin-dependent kinase Cdk2 regulates thymocyte apoptosis
- PMID: 10075979
- PMCID: PMC2193040
- DOI: 10.1084/jem.189.6.957
The cyclin-dependent kinase Cdk2 regulates thymocyte apoptosis
Abstract
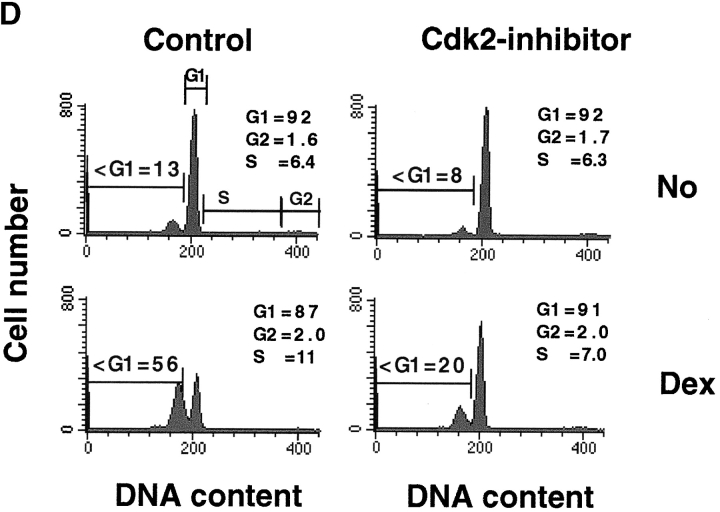
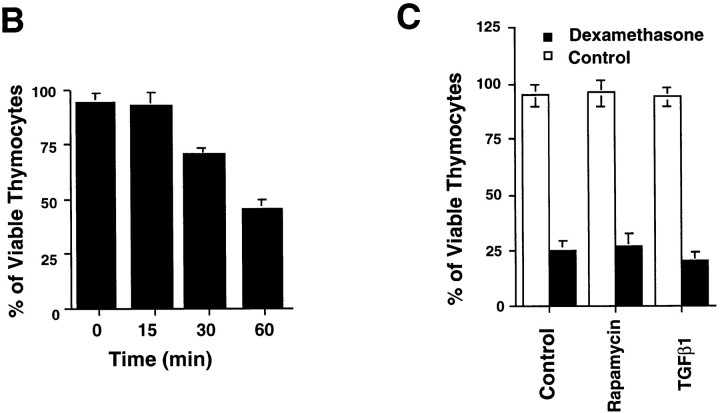
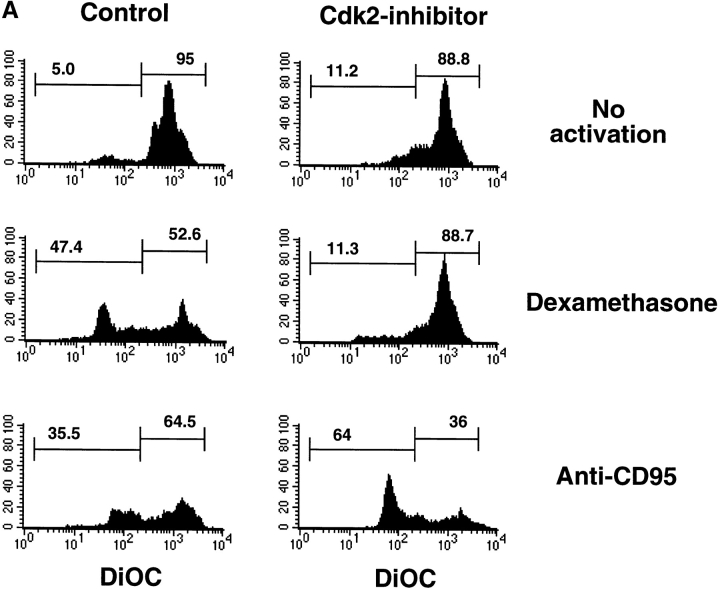
Aberrant activation of cell cycle molecules has been postulated to play a role in apoptosis ("catastrophic cell cycle"). Here we show that in noncycling developing thymocytes, the cyclin- dependent kinase Cdk2 is activated in response to all specific and nonspecific apoptotic stimuli tested, including peptide-specific thymocyte apoptosis. Cdk2 was found to function upstream of the tumor suppressor p53, transactivation of the death promoter Bax, alterations of mitochondrial permeability, Bcl-2, caspase activation, and caspase-dependent proteolytic cleavage of the retinoblastoma protein. Inhibition of Cdk2 completely protected thymocytes from apoptosis, mitochondrial changes, and caspase activation. These data provide the first evidence that Cdk2 activity is crucial for the induction of thymocyte apoptosis.
Figures







Similar articles
-
A link between cell cycle and cell death: Bax and Bcl-2 modulate Cdk2 activation during thymocyte apoptosis.EMBO J. 1998 Dec 15;17(24):7209-18. doi: 10.1093/emboj/17.24.7209. EMBO J. 1998. PMID: 9857178 Free PMC article.
-
Monensin-mediated growth inhibition of SNU-C1 colon cancer cells via cell cycle arrest and apoptosis.Int J Oncol. 2003 Feb;22(2):377-82. Int J Oncol. 2003. PMID: 12527937
-
The subcellular localization of cyclin dependent kinase 2 determines the fate of mesangial cells: role in apoptosis and proliferation.Oncogene. 2002 Mar 7;21(11):1750-8. doi: 10.1038/sj.onc.1205238. Oncogene. 2002. PMID: 11896606
-
How signaling proteins integrate multiple inputs: a comparison of N-WASP and Cdk2.Curr Opin Cell Biol. 2002 Apr;14(2):149-54. doi: 10.1016/s0955-0674(02)00307-1. Curr Opin Cell Biol. 2002. PMID: 11891112 Review.
-
Apoptosis and the cell cycle.Prog Cell Cycle Res. 1996;2:147-63. doi: 10.1007/978-1-4615-5873-6_15. Prog Cell Cycle Res. 1996. PMID: 9552392 Review.
Cited by
-
Mimicking Cdk2 phosphorylation of Bcl-xL at Ser73 results in caspase activation and Bcl-xL cleavage.Cell Death Discov. 2016;2:16001-. doi: 10.1038/cddiscovery.2016.1. Epub 2016 Feb 1. Cell Death Discov. 2016. PMID: 27239332 Free PMC article.
-
Oncogenic and Tumor Suppressive Components of the Cell Cycle in Breast Cancer Progression and Prognosis.Pharmaceutics. 2021 Apr 17;13(4):569. doi: 10.3390/pharmaceutics13040569. Pharmaceutics. 2021. PMID: 33920506 Free PMC article. Review.
-
Caspase-dependent Cdk activity is a requisite effector of apoptotic death events.J Cell Biol. 2000 Jan 10;148(1):59-72. doi: 10.1083/jcb.148.1.59. J Cell Biol. 2000. PMID: 10629218 Free PMC article.
-
Cell cycle and apoptosis.Cell Prolif. 2003 Jun;36(3):165-75. doi: 10.1046/j.1365-2184.2003.00267.x. Cell Prolif. 2003. PMID: 12814432 Free PMC article. Review.
-
Hematopoiesis and thymic apoptosis are not affected by the loss of Cdk2.Mol Cell Biol. 2007 Jul;27(14):5079-89. doi: 10.1128/MCB.00029-07. Epub 2007 May 7. Mol Cell Biol. 2007. PMID: 17485443 Free PMC article.
References
-
- White E. Death-defying acts: a meeting review on apoptosis. Genes Dev. 1993;7:2277–2284. - PubMed
-
- Green DR, Scott DW. Activation-induced apoptosis in lymphocytes. Curr Opin Immunol. 1994;6:476–487. - PubMed
-
- King LB, Ashwell JD. Signaling for death of lymphoid cells. Curr Opin Immunol. 1993;5:368–373. - PubMed
-
- Martin JS, Green RD. Protease activation during apoptosis: death by thousand cuts? . Cell. 1995;82:349–352. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
