

Specific interaction of Tat with the human but not rodent P-TEFb complex mediates the species-specific Tat activation of HIV-1 transcription
- PMID: 10077579
- PMCID: PMC15837
- DOI: 10.1073/pnas.96.6.2728
Specific interaction of Tat with the human but not rodent P-TEFb complex mediates the species-specific Tat activation of HIV-1 transcription
Abstract
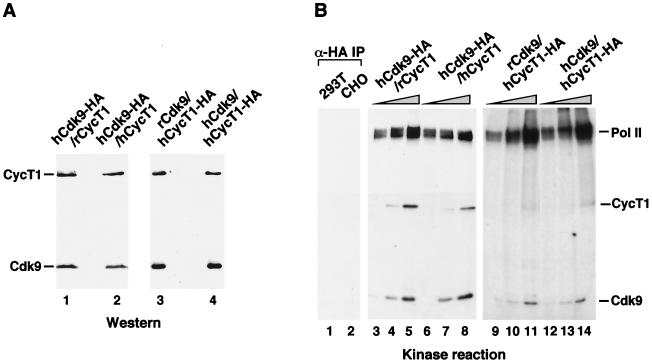
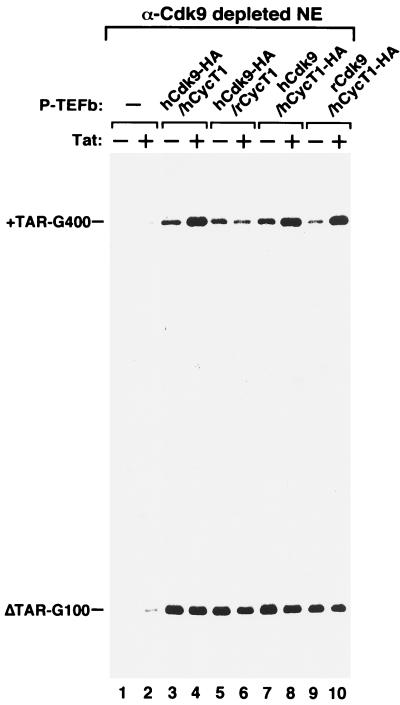
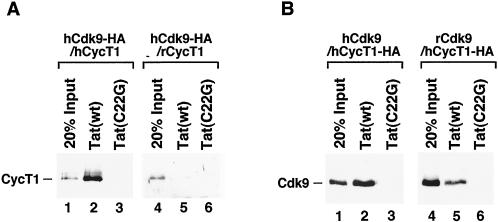
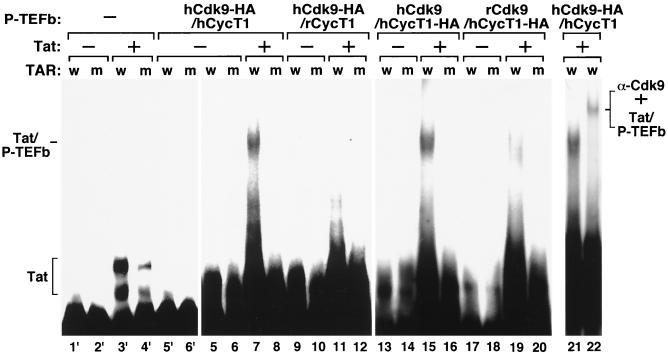
Tat stimulation of HIV-1 transcriptional elongation is species-specific and is believed to require a specific cellular cofactor present in many human and primate cells but not in nonpermissive rodent cells. Human P-TEFb, composed of Cdk9 and cyclin T1, is a general transcription elongation factor that phosphorylates the C-terminal domain of RNA polymerase II. Previous studies have also implicated P-TEFb as a Tat-specific cellular cofactor and, in particular, human cyclin T1 as responsible for the species-specific Tat activation. To obtain functional evidence in support of these hypotheses, we generated and examined the activities of human-rodent "hybrid" P-TEFb complexes. We found that P-TEFb complexes containing human cyclin T1 complexed with either human or rodent Cdk9 supported Tat transactivation and interacted with the Tat activation domain and the HIV-1 TAR RNA element to form TAR loop-dependent ribonucleoprotein complexes. Although a stable complex containing rodent cyclin T1 and human Cdk9 was capable of phosphorylating CTD and mediating basal HIV-1 elongation, it failed to interact with Tat and to mediate Tat transactivation, indicating that the abilities of P-TEFb to support basal elongation and Tat activation can be separated. Together, our data indicated that the specific interaction of human P-TEFb with Tat/TAR, mostly through cyclin T1, is crucial for P-TEFb to mediate a Tat-specific and species-restricted activation of HIV-1 transcription. Amino acid residues unique to human Cdk9 also contributed partially to the formation of the P-TEFb-Tat-TAR complex. Moreover, the cyclin box of cyclin T1 and its immediate flanking region are largely responsible for the specific P-TEFb-Tat interaction.
Figures
Similar articles
-
Relief of two built-In autoinhibitory mechanisms in P-TEFb is required for assembly of a multicomponent transcription elongation complex at the human immunodeficiency virus type 1 promoter.Mol Cell Biol. 2000 Aug;20(16):5897-907. doi: 10.1128/MCB.20.16.5897-5907.2000. Mol Cell Biol. 2000. PMID: 10913173 Free PMC article.
-
Human and rodent transcription elongation factor P-TEFb: interactions with human immunodeficiency virus type 1 tat and carboxy-terminal domain substrate.J Virol. 1999 Jul;73(7):5448-58. doi: 10.1128/JVI.73.7.5448-5458.1999. J Virol. 1999. PMID: 10364292 Free PMC article.
-
Requirement for a kinase-specific chaperone pathway in the production of a Cdk9/cyclin T1 heterodimer responsible for P-TEFb-mediated tat stimulation of HIV-1 transcription.J Biol Chem. 2000 Jan 7;275(1):279-87. doi: 10.1074/jbc.275.1.279. J Biol Chem. 2000. PMID: 10617616
-
Regulatory functions of Cdk9 and of cyclin T1 in HIV tat transactivation pathway gene expression.J Cell Biochem. 1999 Dec 1;75(3):357-68. J Cell Biochem. 1999. PMID: 10536359 Review.
-
Tackling Tat.J Mol Biol. 1999 Oct 22;293(2):235-54. doi: 10.1006/jmbi.1999.3060. J Mol Biol. 1999. PMID: 10550206 Review.
Cited by
-
Spt5 cooperates with human immunodeficiency virus type 1 Tat by preventing premature RNA release at terminator sequences.Mol Cell Biol. 2002 Feb;22(4):1079-93. doi: 10.1128/MCB.22.4.1079-1093.2002. Mol Cell Biol. 2002. PMID: 11809800 Free PMC article.
-
Recruitment of TFIIH to the HIV LTR is a rate-limiting step in the emergence of HIV from latency.EMBO J. 2006 Aug 9;25(15):3596-604. doi: 10.1038/sj.emboj.7601248. Epub 2006 Jul 27. EMBO J. 2006. PMID: 16874302 Free PMC article.
-
Differential acetylation of Tat coordinates its interaction with the co-activators cyclin T1 and PCAF.EMBO J. 2002 Dec 16;21(24):6811-9. doi: 10.1093/emboj/cdf669. EMBO J. 2002. PMID: 12486002 Free PMC article.
-
Tat-SF1 protein associates with RAP30 and human SPT5 proteins.Mol Cell Biol. 1999 Sep;19(9):5960-8. doi: 10.1128/MCB.19.9.5960. Mol Cell Biol. 1999. PMID: 10454543 Free PMC article.
-
Replication of human immunodeficiency viruses engineered with heterologous Tat-transactivation response element interactions.J Virol. 2003 Feb;77(3):1984-91. doi: 10.1128/jvi.77.3.1984-1991.2003. J Virol. 2003. PMID: 12525632 Free PMC article.
References
-
- Cullen B R. Cell. 1993;73:417–420. - PubMed
-
- Jones K A, Peterlin B M. Annu Rev Biochem. 1994;63:717–743. - PubMed
-
- Marciniak R A, Calnan B J, Frankel A D, Sharp P A. Cell. 1990;63:791–802. - PubMed
-
- Sheline C T, Milocco L H, Jones K A. Genes Dev. 1991;5:2508–2520. - PubMed
-
- Selby M J, Peterlin B M. Cell. 1990;62:769–776. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
