

Multiple pathways on a protein-folding energy landscape: kinetic evidence
- PMID: 10077588
- PMCID: PMC15846
- DOI: 10.1073/pnas.96.6.2782
Multiple pathways on a protein-folding energy landscape: kinetic evidence
Abstract
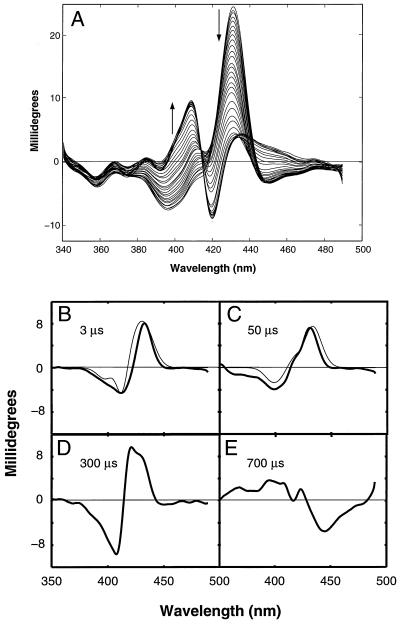
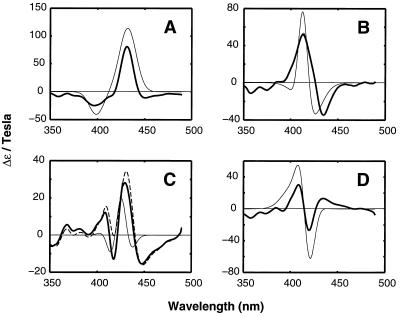
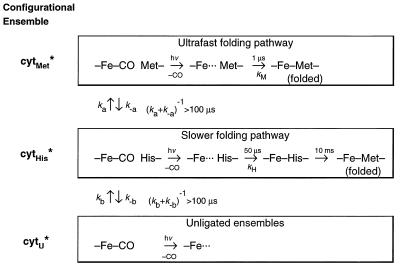
The funnel landscape model predicts that protein folding proceeds through multiple kinetic pathways. Experimental evidence is presented for more than one such pathway in the folding dynamics of a globular protein, cytochrome c. After photodissociation of CO from the partially denatured ferrous protein, fast time-resolved CD spectroscopy shows a submillisecond folding process that is complete in approximately 10(-6) s, concomitant with heme binding of a methionine residue. Kinetic modeling of time-resolved magnetic circular dichroism data further provides strong evidence that a 50-microseconds heme-histidine binding process proceeds in parallel with the faster pathway, implying that Met and His binding occur in different conformational ensembles of the protein, i.e., along respective ultrafast (microseconds) and fast (milliseconds) folding pathways. This kinetic heterogeneity appears to be intrinsic to the diffusional nature of early folding dynamics on the energy landscape, as opposed to the late-time heterogeneity associated with nonnative heme ligation and proline isomers in cytochrome c.
Figures
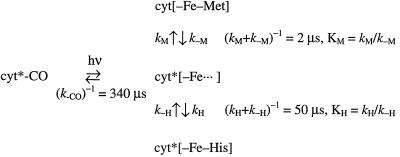
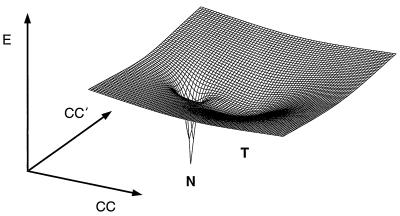
 →hν cyt* + CO, as
a function of protein configurational coordinates (CC). A folding
funnel (N) and an overlapping local minimum (T), corresponding to the
histidine-ligation trap, together form a surface topologically similar
to the “moat” landscape of Dill and Chan (6) describing
heterogeneous (fast and slow) folding kinetics.
→hν cyt* + CO, as
a function of protein configurational coordinates (CC). A folding
funnel (N) and an overlapping local minimum (T), corresponding to the
histidine-ligation trap, together form a surface topologically similar
to the “moat” landscape of Dill and Chan (6) describing
heterogeneous (fast and slow) folding kinetics.References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
