

Angiostatin binds ATP synthase on the surface of human endothelial cells
- PMID: 10077593
- PMCID: PMC15851
- DOI: 10.1073/pnas.96.6.2811
Angiostatin binds ATP synthase on the surface of human endothelial cells
Abstract
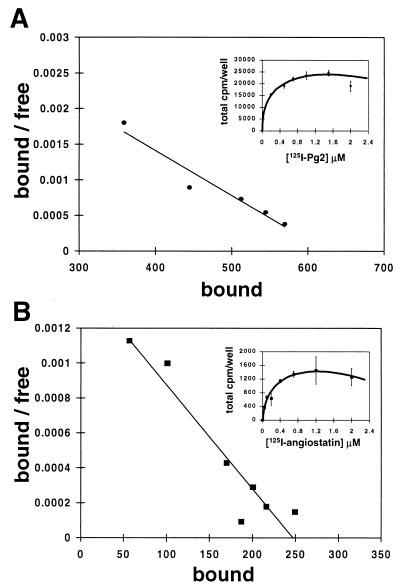
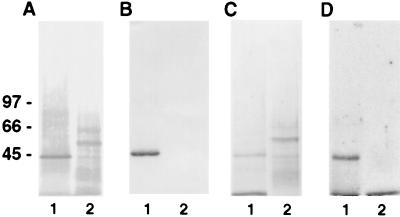
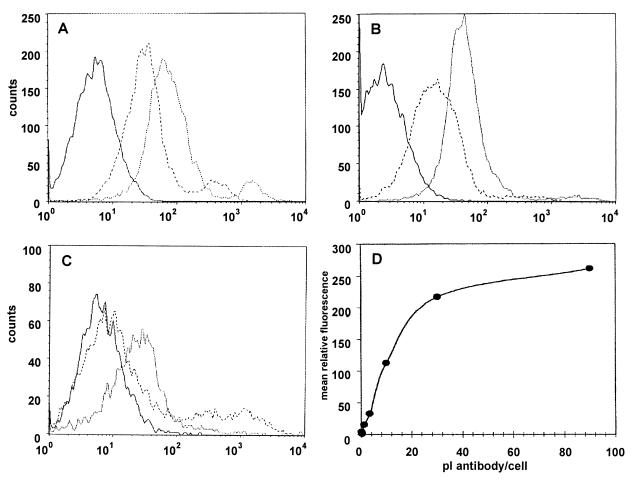
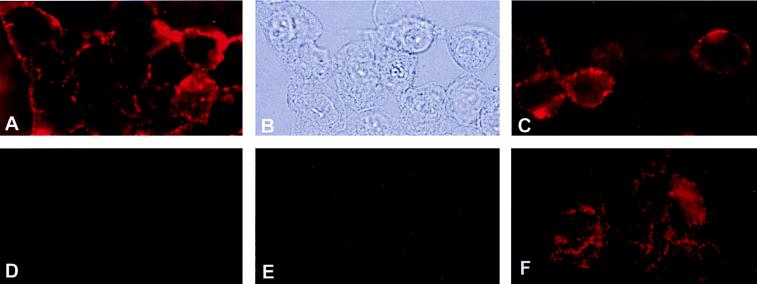
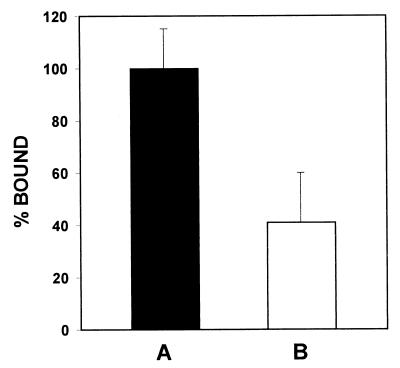
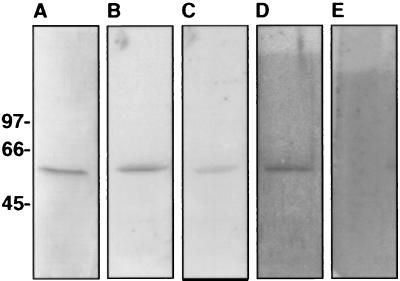
Angiostatin, a proteolytic fragment of plasminogen, is a potent antagonist of angiogenesis and an inhibitor of endothelial cell migration and proliferation. To determine whether the mechanism by which angiostatin inhibits endothelial cell migration and/or proliferation involves binding to cell surface plasminogen receptors, we isolated the binding proteins for plasminogen and angiostatin from human umbilical vein endothelial cells. Binding studies demonstrated that plasminogen and angiostatin bound in a concentration-dependent, saturable manner. Plasminogen binding was unaffected by a 100-fold molar excess of angiostatin, indicating the presence of a distinct angiostatin binding site. This finding was confirmed by ligand blot analysis of isolated human umbilical vein endothelial cell plasma membrane fractions, which demonstrated that plasminogen bound to a 44-kDa protein, whereas angiostatin bound to a 55-kDa species. Amino-terminal sequencing coupled with peptide mass fingerprinting and immunologic analyses identified the plasminogen binding protein as annexin II and the angiostatin binding protein as the alpha/beta-subunits of ATP synthase. The presence of this protein on the cell surface was confirmed by flow cytometry and immunofluorescence analysis. Angiostatin also bound to the recombinant alpha-subunit of human ATP synthase, and this binding was not inhibited by a 2,500-fold molar excess of plasminogen. Angiostatin's antiproliferative effect on endothelial cells was inhibited by as much as 90% in the presence of anti-alpha-subunit ATP synthase antibody. Binding of angiostatin to the alpha/beta-subunits of ATP synthase on the cell surface may mediate its antiangiogenic effects and the down-regulation of endothelial cell proliferation and migration.
Figures

References
-
- O’Reilly M S, Holmgren L, Shing Y, Chen C, Rosenthal R A, Moses M, Lane W S, Cao Y, Sage E H, Folkman J. Cell. 1994;79:315–328. - PubMed
-
- Sottrup-Jensen L, Claeys H, Zajdel M, Petersen T E, Magnusson S. Prog Chem Fibrinolysis Thrombolysis. 1978;3:191–209.
-
- Gately S, Twardowski P, Stack M S, Patrick M, Boggio L, Cundiff D L, Schnaper H W, Madison L, Volpert O, Bouck N, et al. Cancer Res. 1996;36:4887–4890. - PubMed
-
- Dong Z, Kumar R, Yang X, Fidler I J. Cell. 1997;88:801–810. - PubMed
-
- Lijnen H R, Ugwu F, Collen D. Biochemistry. 1998;37:4699–4702. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
