

The role of an alternative sigma factor in motility and pilus formation in the cyanobacterium Synechocystis sp. strain PCC6803
- PMID: 10077659
- PMCID: PMC15917
- DOI: 10.1073/pnas.96.6.3188
The role of an alternative sigma factor in motility and pilus formation in the cyanobacterium Synechocystis sp. strain PCC6803
Abstract
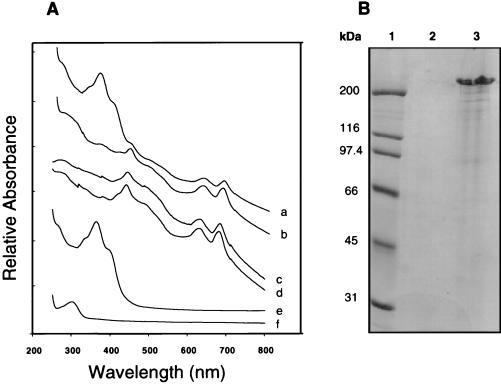
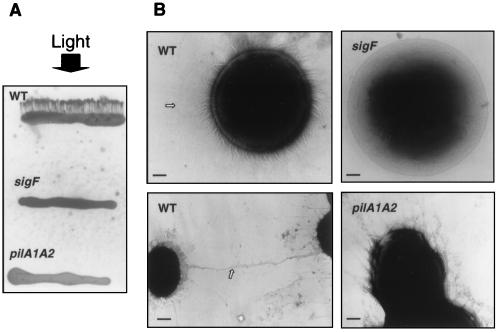
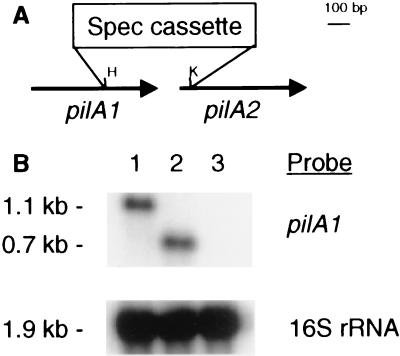
Disruption of a gene for an alternative sigma factor, designated sigF, in the freshwater, unicellular cyanobacterium Synechocystis sp. strain PCC6803 resulted in a pleiotropic phenotype. Most notably, this mutant lost phototactic movement with a concomitant loss of pili, which are abundant on the surface of wild-type cells. The sigF mutant also secreted both high levels of yellow-brown and UV-absorbing pigments and a polypeptide that is similar to a large family of extracellular proteins that includes the hemolysins. Furthermore, the sigF mutant had a dramatically reduced level of the transcript from two tandemly arranged pilA genes (sll1694 and sll1695), which encode major structural components of type IV pili. Inactivation of these pilA genes eliminated phototactic movement, though some pili were still present in this strain. Together, these results demonstrate that SigF plays a critical role in motility via the control of pili formation and is also likely to regulate other features of the cell surface. Furthermore, the data provide evidence that type IV pili are required for phototactic movement in certain cyanobacteria and suggest that different populations of pili present on the Synechocystis cell surface may perform different functions.
Figures

References
-
- Häder D-P. In: The Cyanobacteria. Fay P, Baalen C V, editors. Amsterdam: Elsevier; 1987. pp. 325–346.
-
- Castenholz R W. In: The Biology of Cyanobacteria. Carr N G, Whitton B A, editors. Botanical Monographs 19. Berkeley: Univ. of California Press; 1982. pp. 413–440.
-
- Hoiczyk E, Baumeister W. Mol Microbiol. 1997;26:699–708. - PubMed
-
- Waterbury J B, Willey J M, Franks D G, Valois F W, Watson S W. Science. 1985;230:74–75. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
