

Nucleotide pool imbalance and adenosine deaminase deficiency induce alterations of N-region insertions during V(D)J recombination
- PMID: 10079104
- PMCID: PMC408138
- DOI: 10.1172/JCI4320
Nucleotide pool imbalance and adenosine deaminase deficiency induce alterations of N-region insertions during V(D)J recombination
Abstract
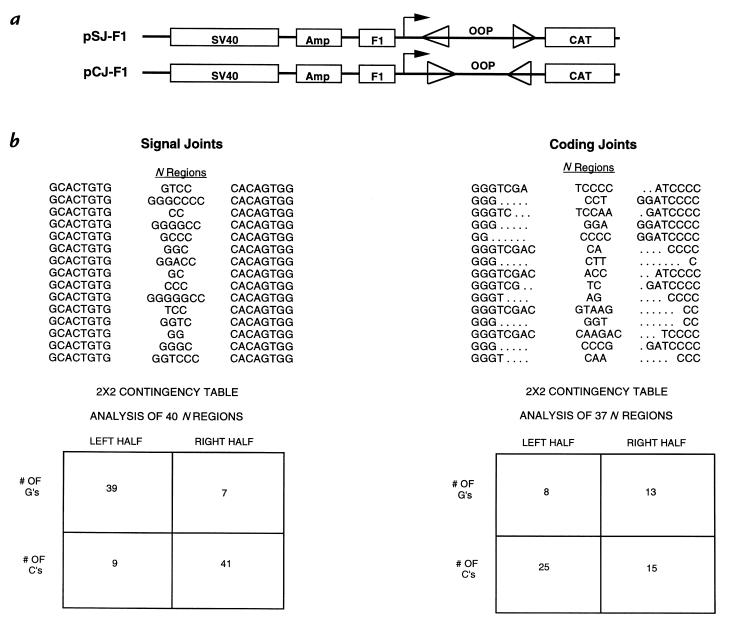
Template-independent nucleotide additions (N regions) generated at sites of V(D)J recombination by terminal deoxynucleotidyl transferase (TdT) increase the diversity of antigen receptors. Two inborn errors of purine metabolism, deficiencies of adenosine deaminase (ADA) and purine nucleoside phosphorylase (PNP), result in defective lymphoid development and aberrant pools of 2'-deoxynucleotides that are substrates for TdT in lymphoid precursors. We have asked whether selective increases in dATP or dGTP pools result in altered N regions in an extrachromosomal substrate transfected into T-cell or pre-B-cell lines. Exposure of the transfected cells to 2'-deoxyadenosine and an ADA inhibitor increased the dATP pool and resulted in a marked increase in A-T insertions at recombination junctions, with an overall decreased frequency of V(D)J recombination. Sequence analysis of VH-DH-JH junctions from the IgM locus in B-cell lines from ADA-deficient patients demonstrated an increase in A-T insertions equivalent to that found in the transfected cells. In contrast, elevation of dGTP pools, as would occur in PNP deficiency, did not alter the already rich G-C content of N regions. We conclude that the frequency of V(D)J recombination and the composition of N-insertions are influenced by increases in dATP levels, potentially leading to alterations in antigen receptors and aberrant lymphoid development. Alterations in N-region insertions may contribute to the B-cell dysfunction associated with ADA deficiency.
Figures


References
-
- Lewis SM. The mechanism of V(D)J joining: lessons from molecular, immunological, and comparative analyses. Adv Immunol. 1994;56:27–150. - PubMed
-
- Thompson CB. New insights into V(D)J recombination and its role in the evolution of the immune system. Immunity. 1995;3:531–539. - PubMed
-
- Ramsden DA, Gellert M. Formation and resolution of double-strand break intermediates in V(D)J rearrangement. Genes Dev. 1995;9:2409–2420. - PubMed
-
- Gilfillan S, Dierich A, Lemeur M, Benoist C, Mathis D. Mice lacking TdT: mature animals with an immature lymphocyte repertoire. Science. 1993;261:1175–1178. - PubMed
-
- Komori T, Okada A, Stewart V, Alt FW. Lack of N regions in antigen receptor variable genes of TdT-deficient lymphocytes. Science. 1993;261:1171–1174. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
