

Targeted mutation of plasma phospholipid transfer protein gene markedly reduces high-density lipoprotein levels
- PMID: 10079112
- PMCID: PMC408146
- DOI: 10.1172/JCI5578
Targeted mutation of plasma phospholipid transfer protein gene markedly reduces high-density lipoprotein levels
Abstract
It has been proposed that the plasma phospholipid transfer protein (PLTP) facilitates the transfer of phospholipids and cholesterol from triglyceride-rich lipoproteins (TRL) into high-density lipoproteins (HDL). To evaluate the in vivo role of PLTP in lipoprotein metabolism, we used homologous recombination in embryonic stem cells and produced mice with no PLTP gene expression. Analysis of plasma of F2 homozygous PLTP-/- mice showed complete loss of phosphatidylcholine, phosphatidylethanolamine, phosphatidylinositol, sphingomyelin, and partial loss of free cholesterol transfer activities. Moreover, the in vivo transfer of [3H]phosphatidylcholine ether from very-low-density proteins (VLDL) to HDL was abolished in PLTP-/- mice. On a chow diet, PLTP-/- mice showed marked decreases in HDL phospholipid (60%), cholesterol (65%), and apo AI (85%), but no significant change in non-HDL lipid or apo B levels, compared with wild-type littermates. On a high-fat diet, HDL levels were similarly decreased, but there was also an increase in VLDL and LDL phospholipids (210%), free cholesterol (60%), and cholesteryl ester (40%) without change in apo B levels, suggesting accumulation of surface components of TRL. Vesicular lipoproteins were shown by negative-stain electron microscopy of the free cholesterol- and phospholipid-enriched IDL/LDL fraction. Thus, PLTP is the major factor facilitating transfer of VLDL phospholipid into HDL. Reduced plasma PLTP activity causes markedly decreased HDL lipid and apoprotein, demonstrating the importance of transfer of surface components of TRL in the maintenance of HDL levels. Vesicular lipoproteins accumulating in PLTP-/- mice on a high-fat diet could influence the development of atherosclerosis.
Figures




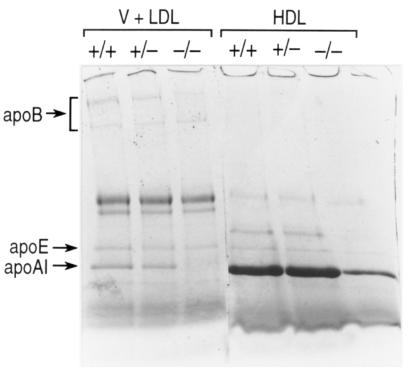

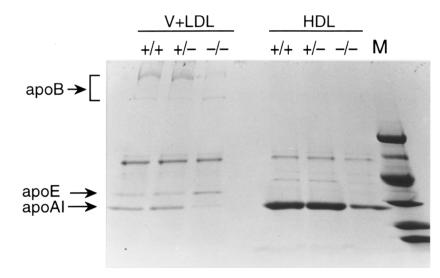

References
-
- Rhoads GG, Gulbrandsen CL, Kagan A. Serum lipoproteins and coronary heart disease in a population study of Hawaii Japanese men. N Engl J Med. 1976;294:293–298. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
