

Review
doi: 10.1016/s0002-9440(10)65311-4.
Rous-Whipple Award Lecture. Chemical features of peptide selection by the class II histocompatibility molecules
Affiliations
- PMID: 10079241
- PMCID: PMC1866414
- DOI: 10.1016/s0002-9440(10)65311-4
Item in Clipboard
Review
Rous-Whipple Award Lecture. Chemical features of peptide selection by the class II histocompatibility molecules
Am J Pathol.
1999 Mar.
No abstract available
Figures
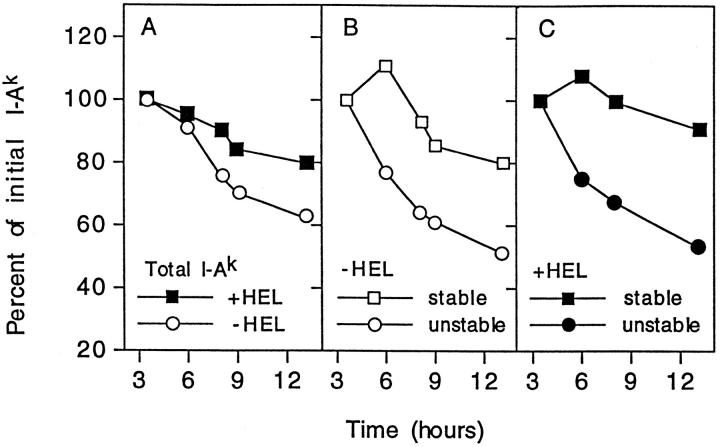
The time of persistence of class II MHC molecules depends on their content of strong or weak binding peptides. The C3F6 line was pulsed with radioactive amino acid precursors, then chased. At each time point the radioactivity associated with total class II MHC molecules and with the two SDS subsets was determined. The T 1/2 calculated by linear regression analysis in cells cultured without HEL was 12.6 hours: 10.2 hours for the unstable SDS set and 20.3 hours for the stable set. In cells cultured with HEL, the T 1/2 was 23 hours, with 10.9 hours for the unstable and 43 hours for the stable SDS set. Reprinted with permission from Nature 371:250–252, copyright 1994 Macmillan Magazines Limited.
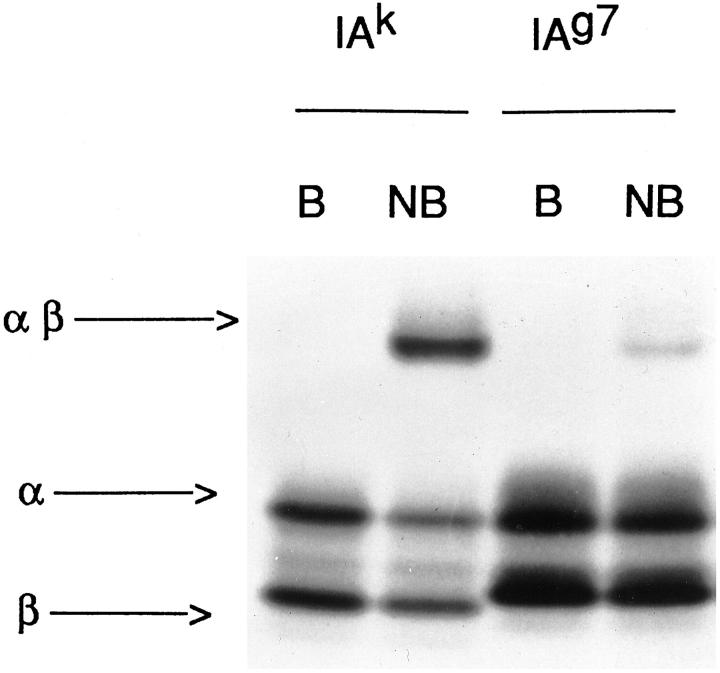
Two sets of class II MHC molecules can be distinguished by their migration in SDS-PAGE when the molecules are not boiled (NB). After 100°C (B), the molecule dissociates in the α and β chains. The I-Ak molecules contain about 50% stable, which contain the strong binding peptides. The SDS-unstable are made up of weakly binding peptides. The I-Ag7 molecule bind peptides weakly and are mostly found in SDS-unstable molecules.
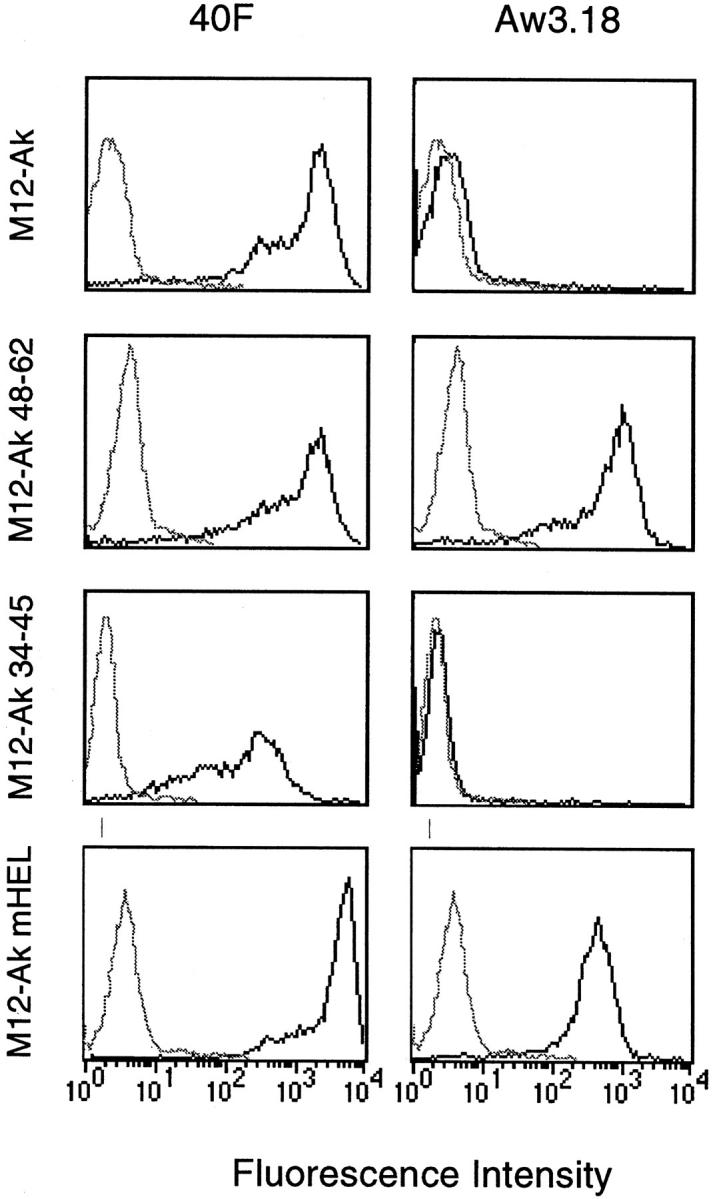
A monoclonal antibody can be made that specifically recognizes the I-Ak−48–62 complex found in APC fed HEL. The panels to the left represent cells reacted with a monoclonal antibody (40F) that recognizes all I-Ak molecules. On the right is the specific monoclonal antibody Aw3.18. M12-Ak refers to the C3.F6 line bearing I-Ak, not exposed to HEL: the 40F-specific (bold line) can be seen, but there is no reactivity with Aw3.18. M12-Ak−48–61 is a line engineered to express Aβk chains with covalently linked 48–62 peptide sequence: these APC react with Aw3.18. M12-Ak−34–45 is the line containing an unrelated complex, indicating no reactivity with Aw3.18. Lastly, M12-Ak-mHEL is a line bearing HEL molecules, known to donate 48–62 peptide to their I-Ak molecules; these peptides are identified with Aw3.18.
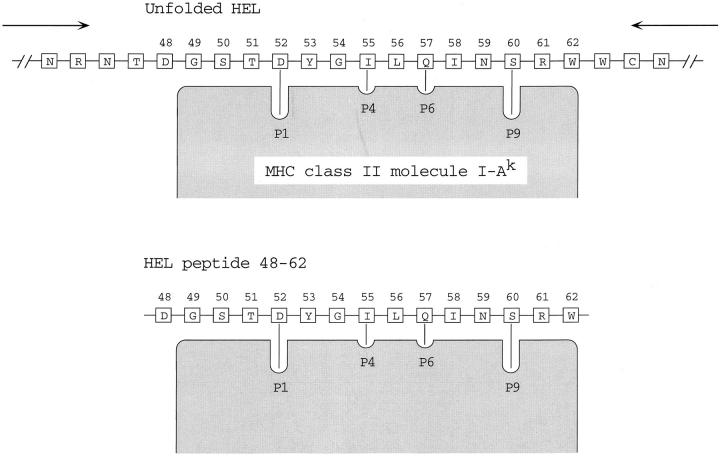
HEL is unfolded and binds to I-Ak through its 52–61 core segment. After binding, the unfolded polypeptide is trimmed up to the edge of the combining site.
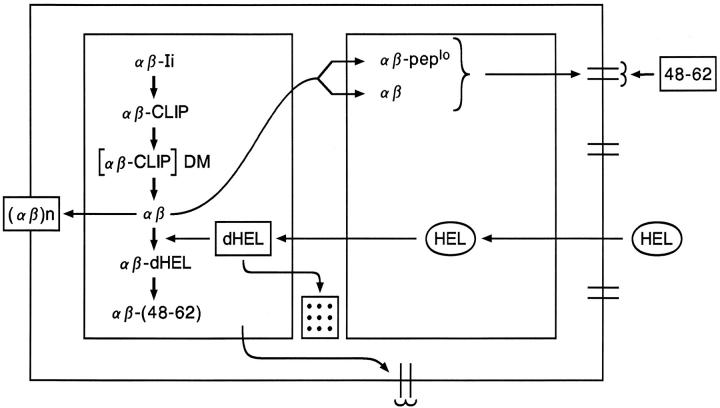
A scenario for the processing of HEL in APC. The description is in the text.
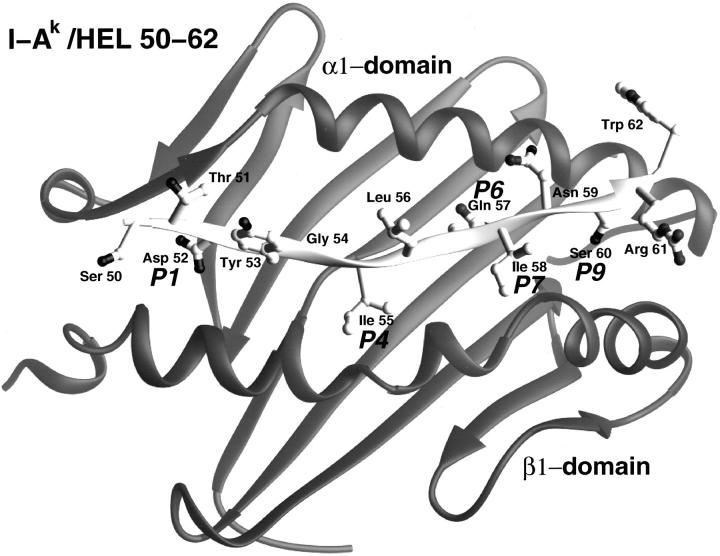
Ribbon diagram of the I-Ak−48–62 complex; taken from the X-ray crystal structure by D. Fremont. The important P1 pocket is filled by the side chain of the aspartic acid (Asp 52). The TcR contacts are Tyr 53, Leu 56, Asn 59, and Trp 62.
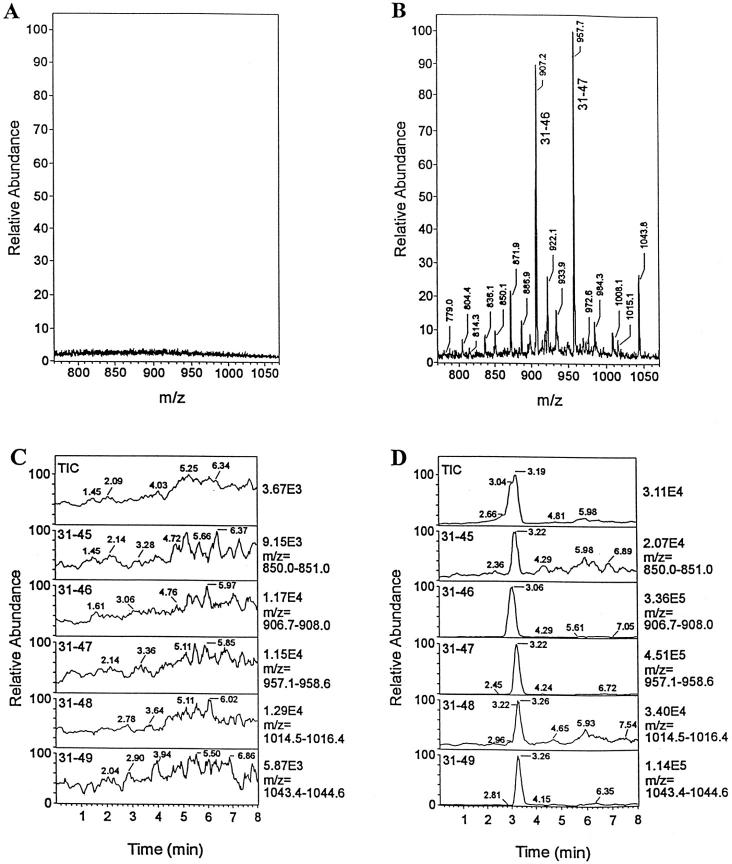
Identification of minor HEL peptides by immunoaffinity capture. The peptides bound to I-Ak from APC either not fed, or fed, with HEL were isolated and passed through a column containing an antibody to the 34–45 sequence. The peptides were then released and examined by tandem mass spectrometry. No peptides were identified from the column to which the I-Ak extract from APC not cultured with HEL (A and C). A series of peptides could be identified from the 31–49 family; the precise sequence was then identified. Reprinted with permission from ref 65, copyright 1998 American Association of Immunologists.
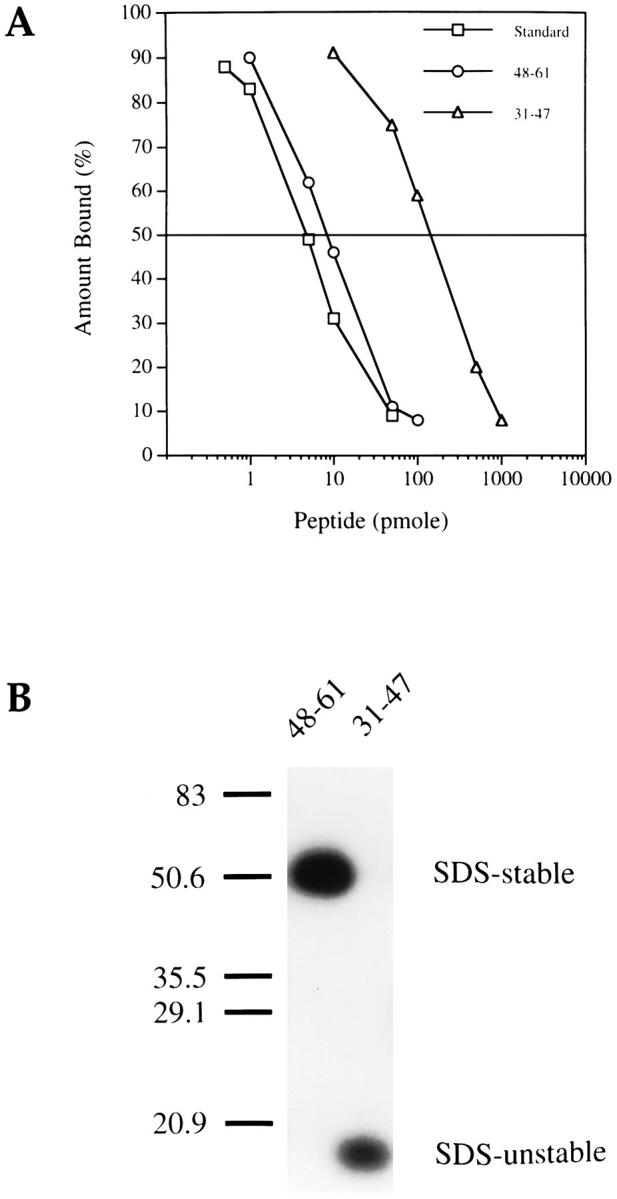
The peptides identified in Figure 7 ▶ are weak binders to I-Ak. These two figures compare the binding to I-Ak of the dominant 48–61 and minor 31–47 HEL determinant (A): about 30-fold more of the latter is required to inhibit binding to the standard peptide. B shows that the strong complex of I-Ak−48–61, but not that with 31–47, is resistant to SDS-PAGE. Reprinted with permission from ref 65, copyright 1998 American Association of Immunologists.
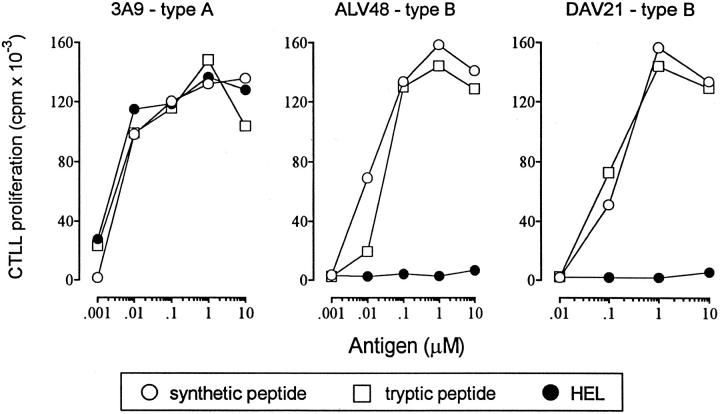
Two sets of 48–62-reactive T cells can be identified: type A and type B. Each set was tested on HEL, a tryptic peptide from residues 46 to 61, and with a 48–61 synthetic peptide. Type A T cells are induced by immunization with HEL, react with 48–62 derived from intracellular processing and from 48–61 offered to the cell as either a synthetic or a tryptic peptide. Type B T cells are induced by immunization with the peptide and react only with the peptide, not with the 48–62 derived from processing. Reprinted with permission from J Immunol 156:2365–2368, copyright 1996 American Association of Immunologists.
Similar articles
-
Rules for peptide presentation by MHC class II molecules.Int Rev Immunol. 1993;10(4):301-11. doi: 10.3109/08830189309061705. Int Rev Immunol. 1993. PMID: 8294843 Review. No abstract available.
-
Structure of peptides associated with class I and class II MHC molecules.Annu Rev Immunol. 1994;12:181-207. doi: 10.1146/annurev.iy.12.040194.001145. Annu Rev Immunol. 1994. PMID: 7516668 Review.
-
Interactions between peptides and major histocompatibility complex molecules during antigen processing and presentation.Chem Immunol. 1993;57:63-87. Chem Immunol. 1993. PMID: 8260090 Review. No abstract available.
-
T-splenocytes from non-obese diabetic mice binding to xenogeneic pancreatic beta-cells in vitro. Implication of the alpha/beta T-cell receptor and of major histocompatibility complex class II molecules from target cells.J Autoimmun. 1993 Dec;6(6):753-69. doi: 10.1006/jaut.1993.1062. J Autoimmun. 1993. PMID: 8155255
-
Antigen presentation in the autoimmune diabetes of the NOD mouse.Annu Rev Immunol. 2014;32:579-608. doi: 10.1146/annurev-immunol-032712-095941. Epub 2014 Feb 5. Annu Rev Immunol. 2014. PMID: 24499272 Review.
References
-
- Unanue ER, Allen PM: The basis for the immunoregulatory role of macrophages and other accessory cells. Science 1987, 236:551-557 - PubMed
-
- Unanue ER: Studies in Listeriosis show the strong symbiosis between the innate cellular system and the T cell response. Immunol Rev 1997, 158:11-25 - PubMed
-
- Lenschow DJ, Walunas TL, Bluestone JA: CD28/B7 system of T cell costimulation Annu Rev Immunol 1996, 14:233-258 - PubMed
-
- Hsieh CS, Macatonia SE, Tripp CS, Wolf SF, O’Garra A, Murphy SM: Development of Th1 CD4+ T cells through IL-12 produced by Listeria-induced macrophages. Science 1993, 260:547-549 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
