

Tat activates human immunodeficiency virus type 1 transcriptional elongation independent of TFIIH kinase
- PMID: 10082552
- PMCID: PMC84079
- DOI: 10.1128/MCB.19.4.2863
Tat activates human immunodeficiency virus type 1 transcriptional elongation independent of TFIIH kinase
Abstract
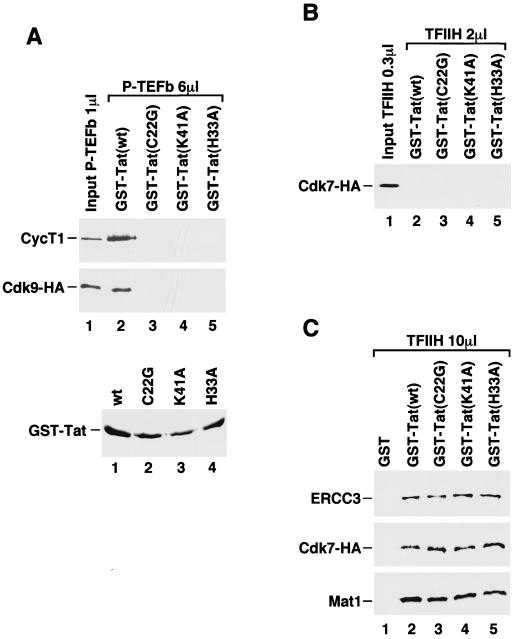
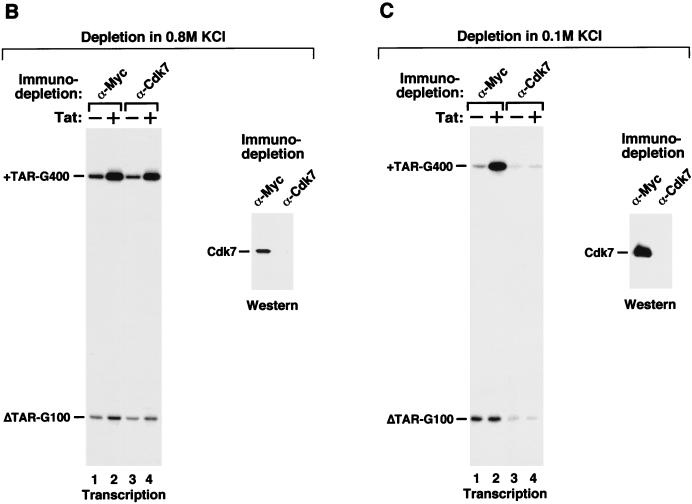
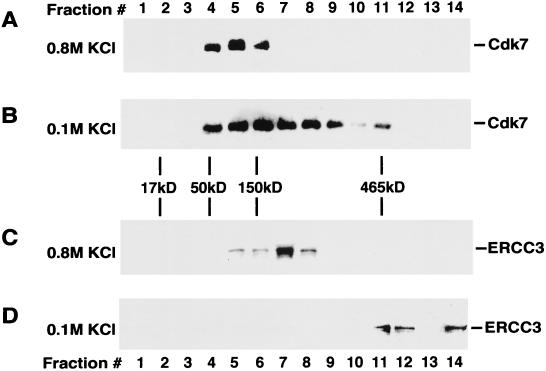
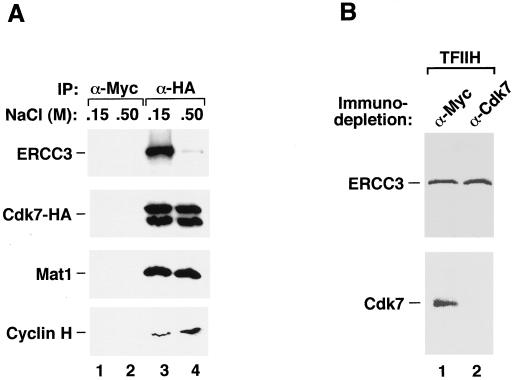
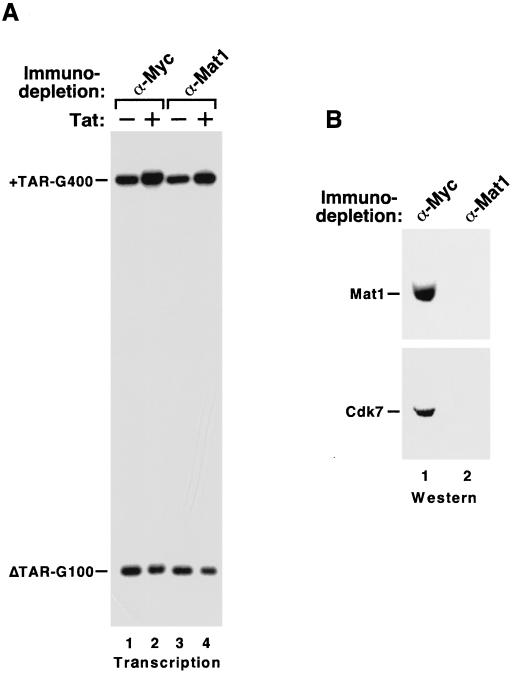
Tat stimulates human immunodeficiency virus type 1 (HIV-1) transcriptional elongation by recruitment of the human transcription elongation factor P-TEFb, consisting of Cdk9 and cyclin T1, to the HIV-1 promoter via cooperative binding to the nascent HIV-1 transactivation response RNA element. The Cdk9 kinase activity has been shown to be essential for P-TEFb to hyperphosphorylate the carboxy-terminal domain (CTD) of RNA polymerase II and mediate Tat transactivation. Recent reports have shown that Tat can also interact with the multisubunit transcription factor TFIIH complex and increase the phosphorylation of CTD by the Cdk-activating kinase (CAK) complex associated with the core TFIIH. These observations have led to the proposal that TFIIH and P-TEFb may act sequentially and in a concerted manner to promote phosphorylation of CTD and increase polymerase processivity. Here, we show that under conditions in which a specific and efficient interaction between Tat and P-TEFb is observed, only a weak interaction between Tat and TFIIH that is independent of critical amino acid residues in the Tat transactivation domain can be detected. Furthermore, immunodepletion of CAK under high-salt conditions, which allow CAK to be dissociated from core-TFIIH, has no effect on either basal HIV-1 transcription or Tat activation of polymerase elongation in vitro. Therefore, unlike the P-TEFb kinase activity that is essential for Tat activation of HIV-1 transcriptional elongation, the CAK kinase associated with TFIIH appears to be dispensable for Tat function.
Figures

References
-
- Akoulitchev S, Makela T P, Weinberg R A, Reinberg D. Requirement for TFIIH kinase activity in transcription by RNA polymerase II. Nature. 1995;377:557–560. - PubMed
-
- Chun R F, Jeang K T. Requirements for RNA polymerase II carboxyl-terminal domain for activated transcription of human retroviruses human T-cell lymphotropic virus I and HIV-1. J Biol Chem. 1996;271:27888–27894. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous