

Hypersensitive site 2 specifies a unique function within the human beta-globin locus control region to stimulate globin gene transcription
- PMID: 10082573
- PMCID: PMC84100
- DOI: 10.1128/MCB.19.4.3062
Hypersensitive site 2 specifies a unique function within the human beta-globin locus control region to stimulate globin gene transcription
Abstract
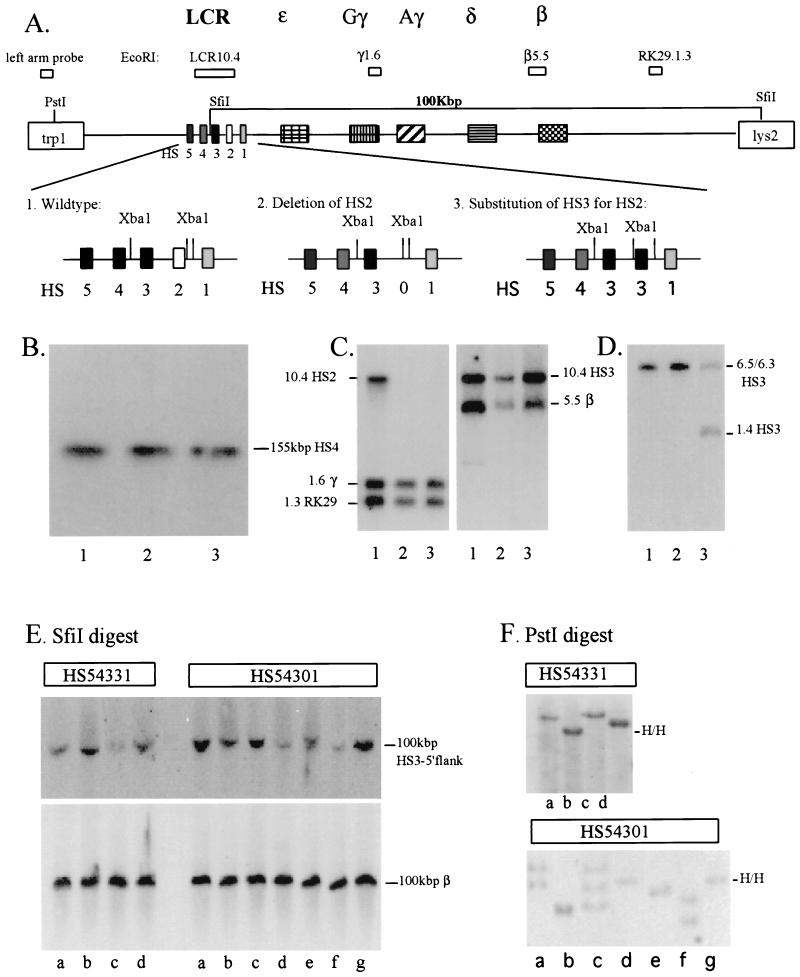
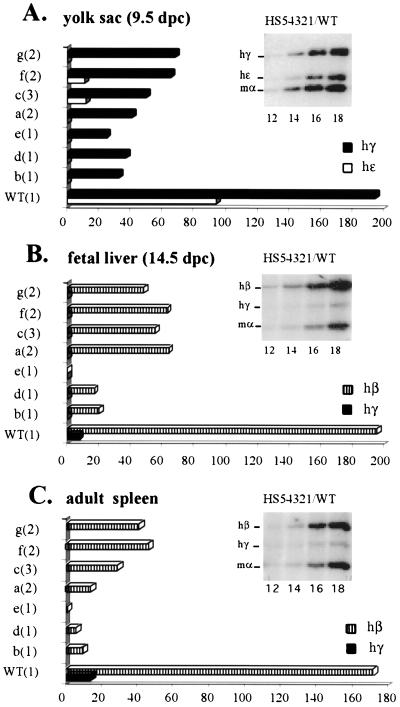
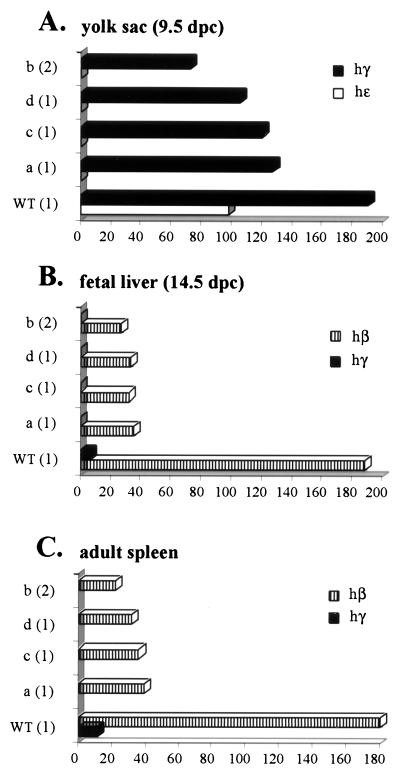
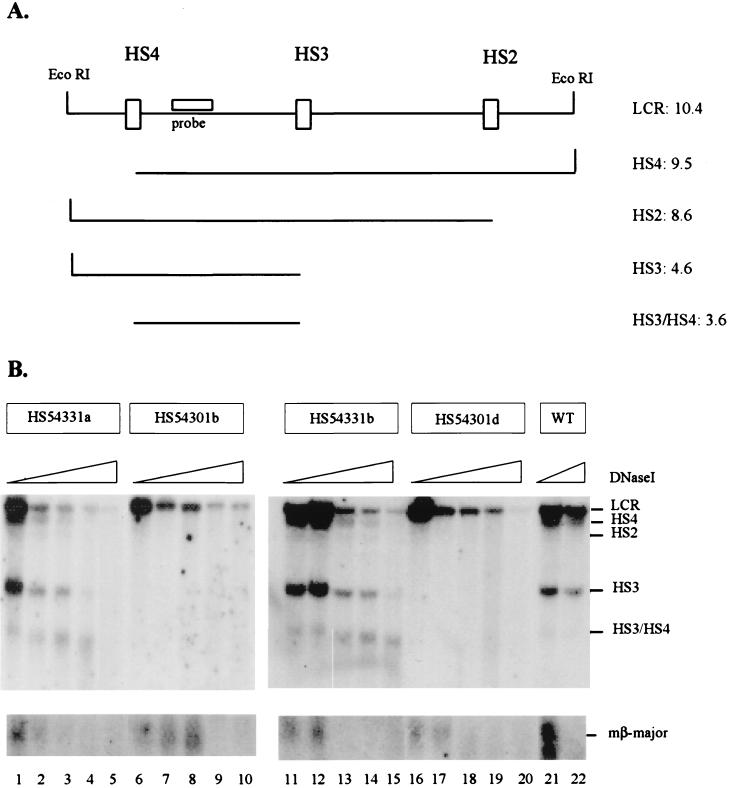
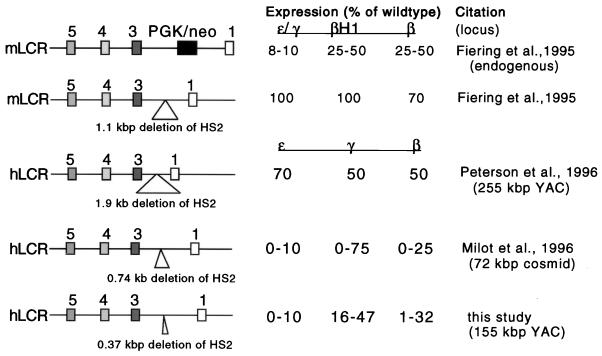
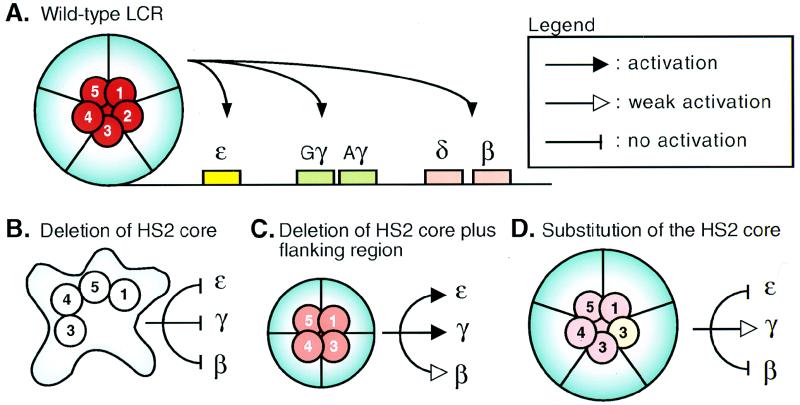
The human beta-globin locus control region (LCR) harbors both strong chromatin opening and enhancer activity when assayed in transgenic mice. To understand the contribution of individual DNase I hypersensitive sites (HS) to the function of the human beta-globin LCR, we have mutated the core elements within the context of a yeast artificial chromosome (YAC) carrying the entire locus and then analyzed the effect of these mutations on the formation of LCR HS elements and expression of the genes in transgenic mice. In the present study, we examined the consequences of two different HS2 mutations. We first generated seven YAC transgenic lines bearing a deletion of the 375-bp core enhancer of HS2. Single-copy HS2 deletion mutants exhibited severely depressed HS site formation and expression of all of the human beta-globin genes at every developmental stage, confirming that HS2 is a vital, integral component of the LCR. We also analyzed four transgenic lines in which the core element of HS2 was replaced by that of HS3 and found that while HS3 is able to restore the chromatin-opening activity of the LCR, it is not able to functionally replace HS2 in mediating high-level globin gene transcription. These results continue to support the hypothesis that HS2, HS3, and HS4 act as a single, integral unit to regulate human globin gene transcription as a holocomplex, but they can also be interpreted to say that formation of a DNase I hypersensitive holocomplex alone is not sufficient for mediating high-level globin gene transcription. We therefore propose that the core elements must productively interact with one another to generate a unique subdomain within the nucleoprotein holocomplex that interacts in a stage-specific manner with individual globin gene promoters.
Figures
References
-
- Aladjem M I, Groudine M, Brody L L, Dieken E S, Fournier R E, Wahl G M, Epner E M. Participation of the human β-globin locus control region in initiation of DNA replication. Science. 1995;270:815–819. - PubMed
-
- Bungert J, Dave U, Lieuw K E, Lim K-C, Shavit J A, Liu Q, Engel J D. Synergistic regulation of human β-globin gene switching by locus control region elements HS3 and HS4. Genes Dev. 1995;9:3083–3096. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources