

Calreticulin is essential for cardiac development
- PMID: 10085286
- PMCID: PMC2148186
- DOI: 10.1083/jcb.144.5.857
Calreticulin is essential for cardiac development
Abstract
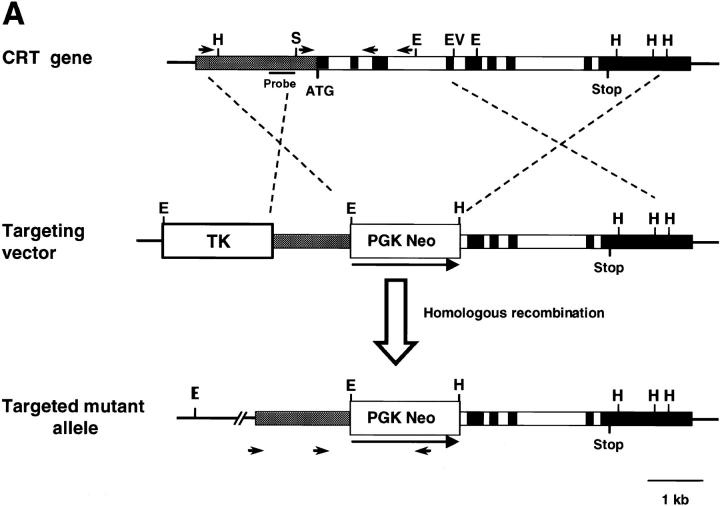
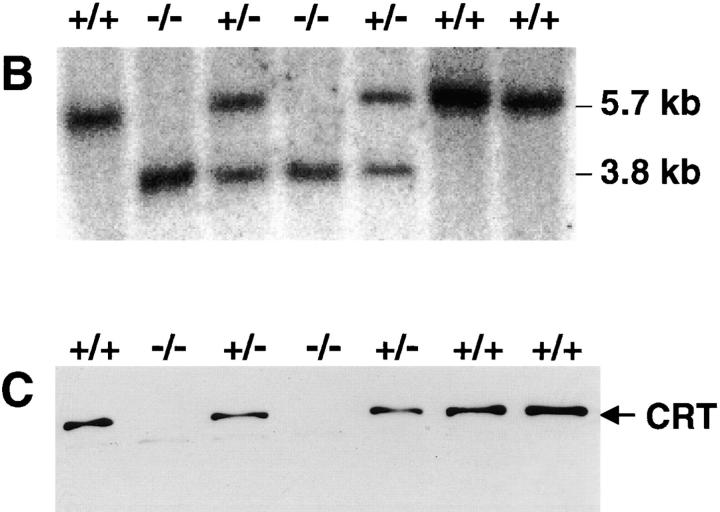
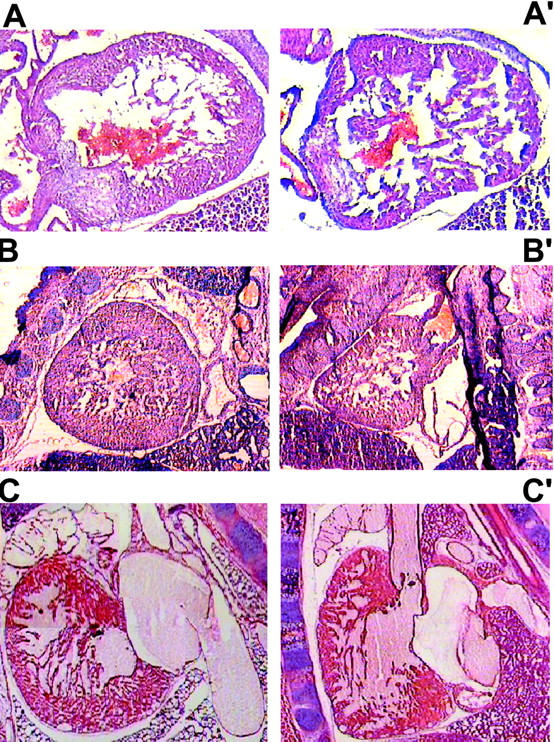
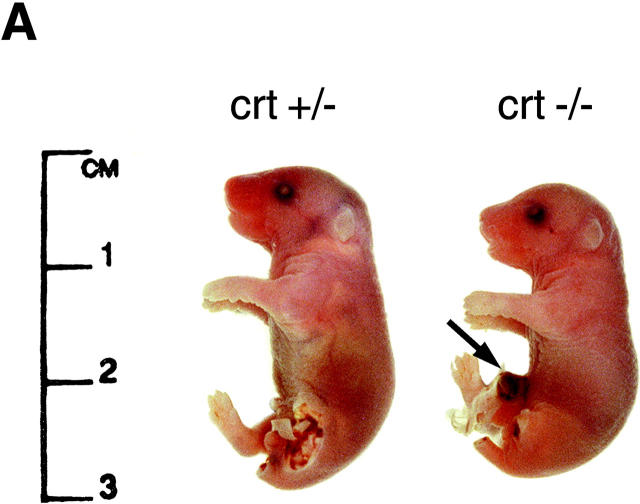
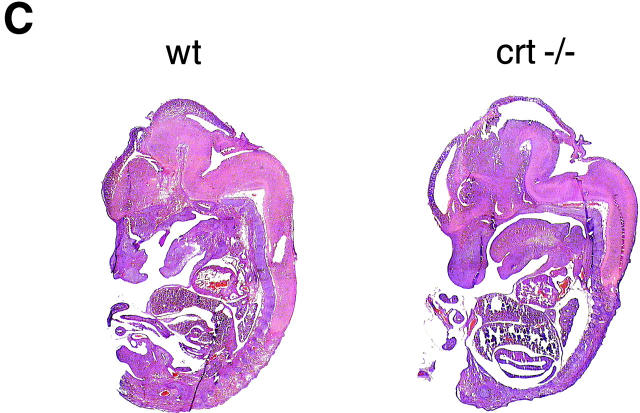
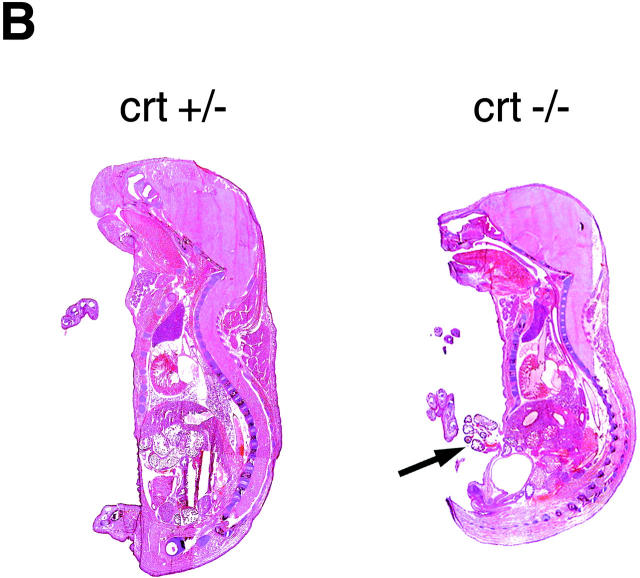
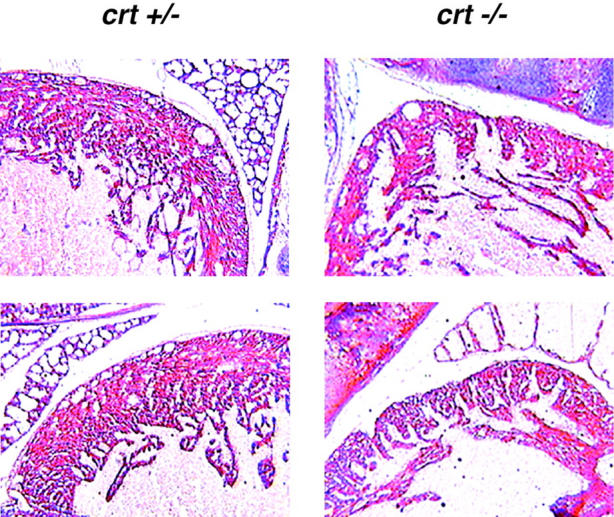
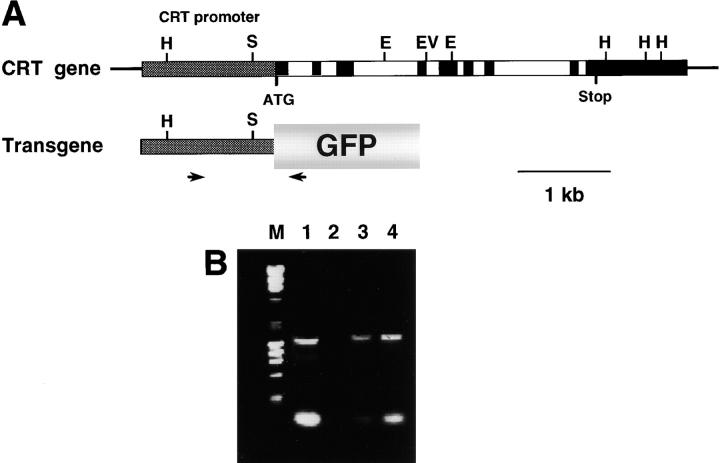
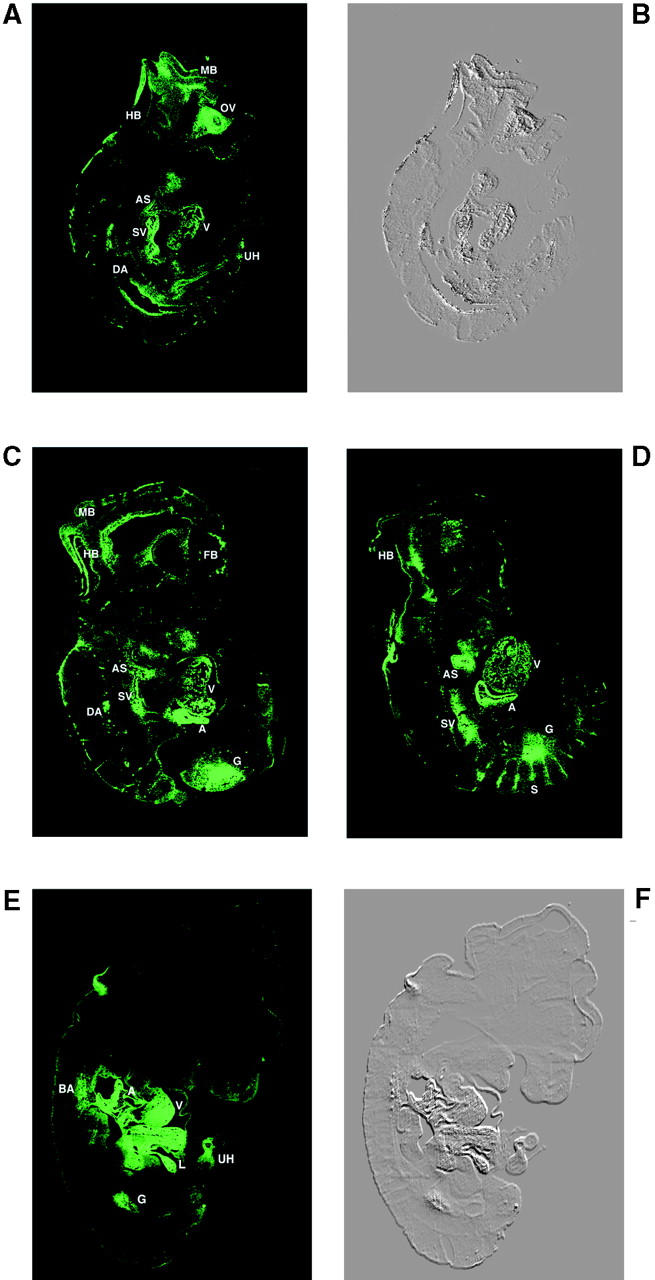
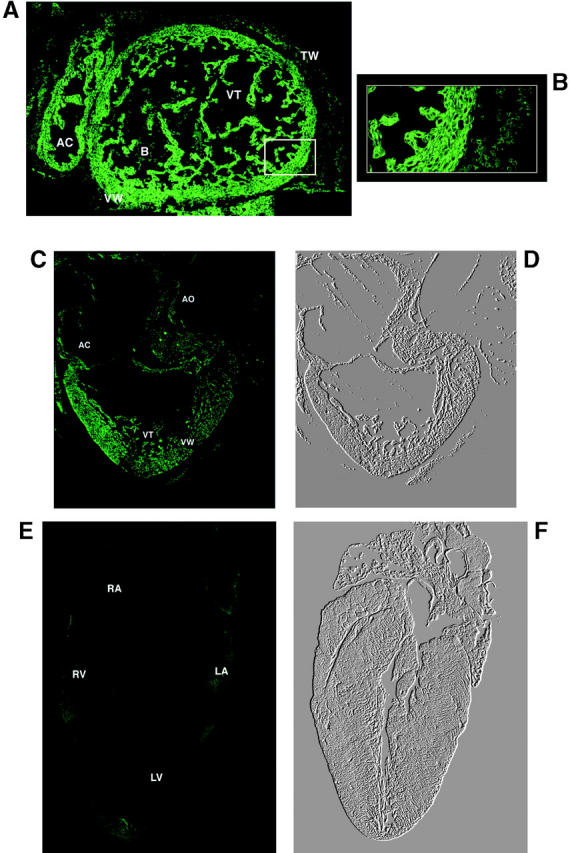
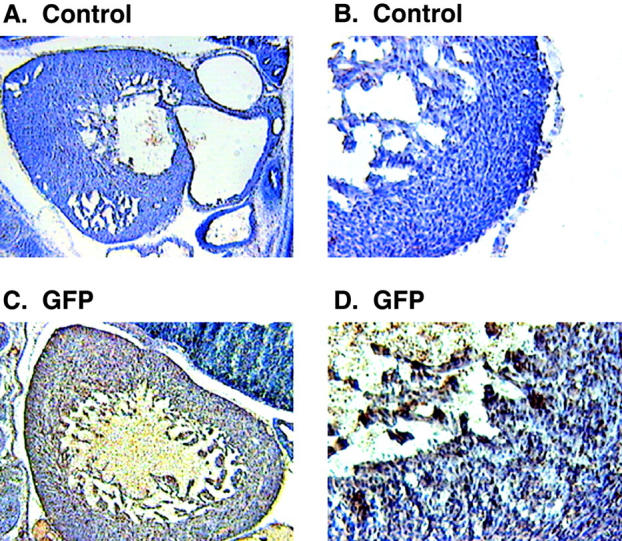
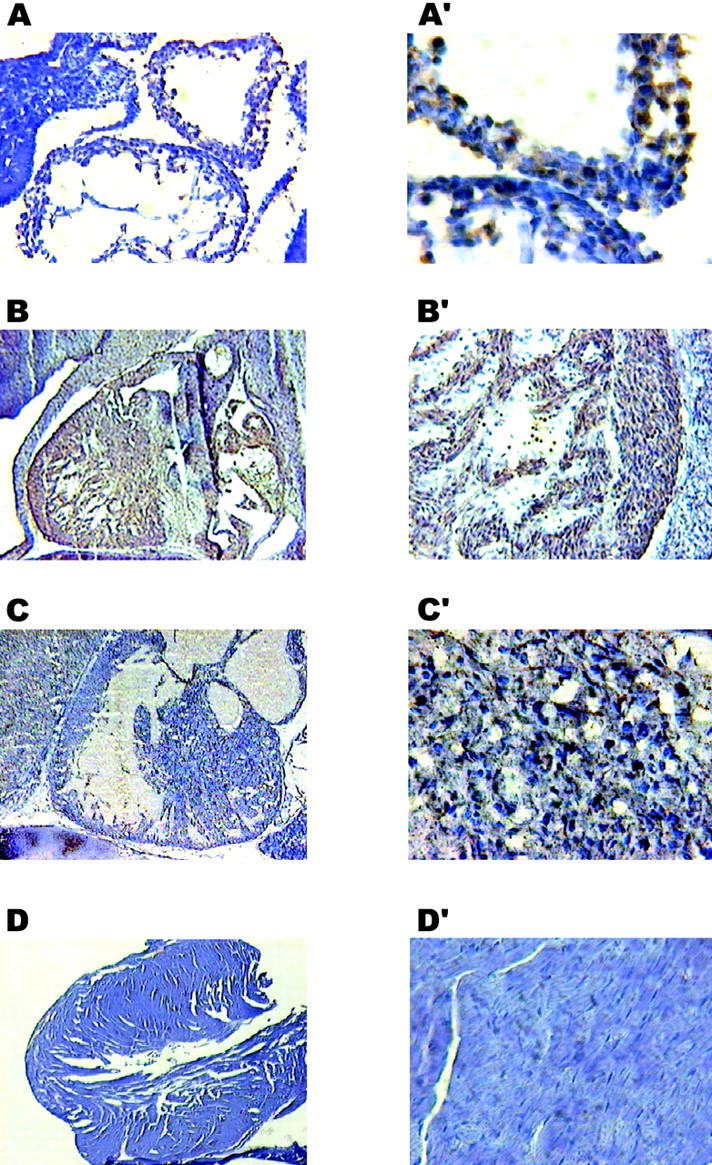
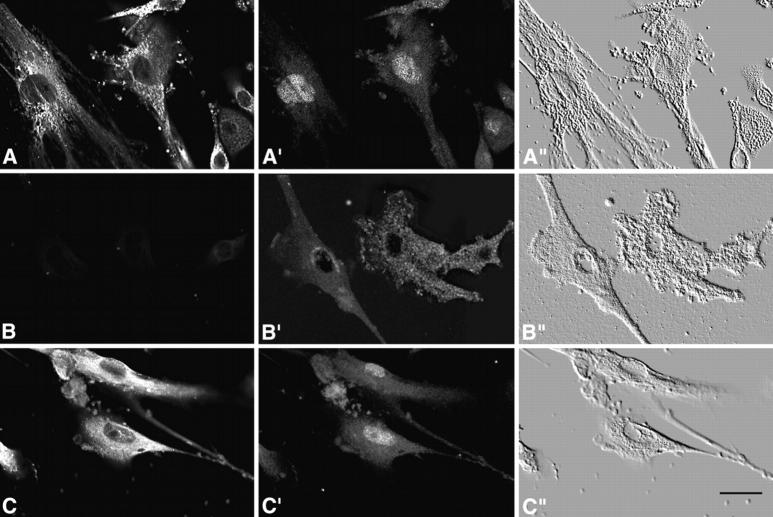
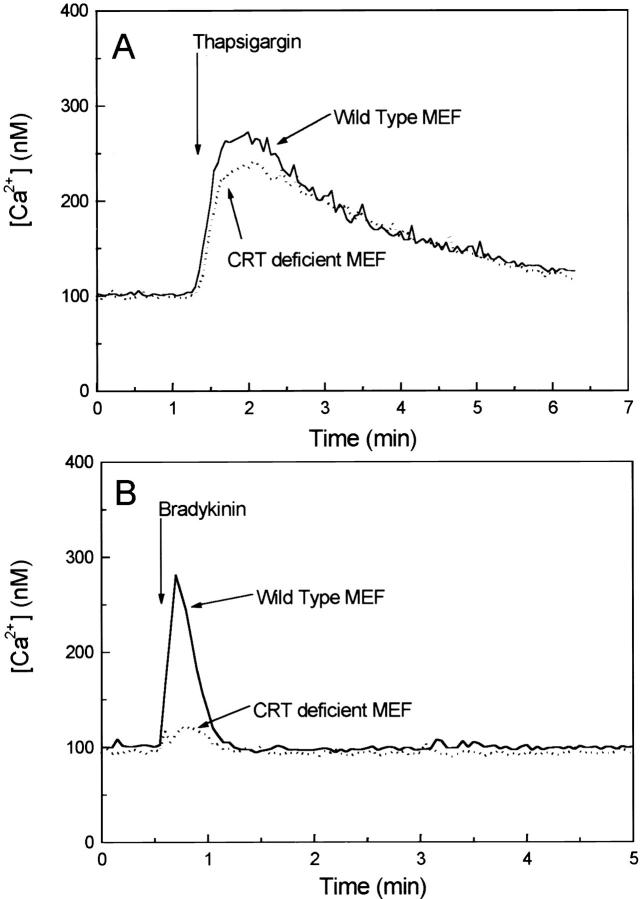
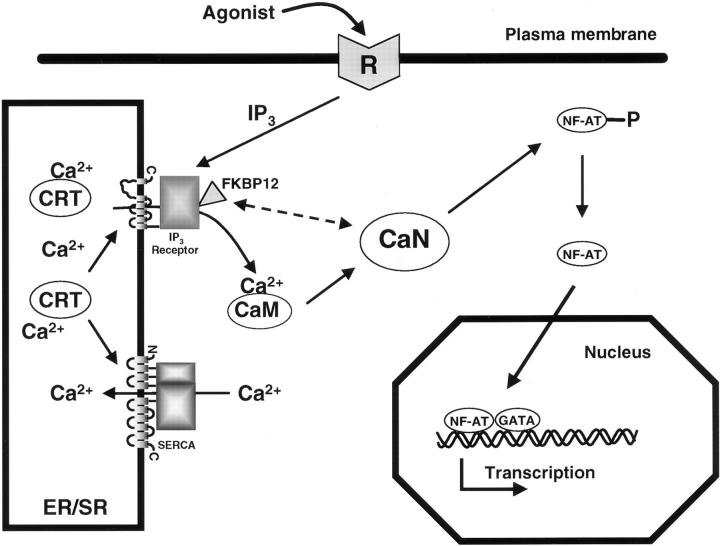
Calreticulin is a ubiquitous Ca2+ binding protein, located in the endoplasmic reticulum lumen, which has been implicated in many diverse functions including: regulation of intracellular Ca2+ homeostasis, chaperone activity, steroid-mediated gene regulation, and cell adhesion. To understand the physiological function of calreticulin we used gene targeting to create a knockout mouse for calreticulin. Mice homozygous for the calreticulin gene disruption developed omphalocele (failure of absorption of the umbilical hernia) and showed a marked decrease in ventricular wall thickness and deep intertrabecular recesses in the ventricular walls. Transgenic mice expressing a green fluorescent protein reporter gene under the control of the calreticulin promoter were used to show that the calreticulin gene is highly activated in the cardiovascular system during the early stages of cardiac development. Calreticulin protein is also highly expressed in the developing heart, but it is only a minor component of the mature heart. Bradykinin-induced Ca2+ release by the InsP3-dependent pathway was inhibited in crt-/- cells, suggesting that calreticulin plays a role in Ca2+ homeostasis. Calreticulin-deficient cells also exhibited impaired nuclear import of nuclear factor of activated T cell (NF-AT3) transcription factor indicating that calreticulin plays a role in cardiac development as a component of the Ca2+/calcineurin/NF-AT/GATA-4 transcription pathway.
Figures
References
-
- Andrin C, Pinkoski MJ, Burns K, Atkinson EA, Krahenbuhl O, Hudig D, Fraser SA, Winkler U, Tschopp J, Opas M, et al. Interaction between a Ca2+binding protein calreticulin and perforin, a component of the T-cell granules. Biochemistry. 1998;37:10386–10394. - PubMed
-
- Ausubel, F.M., R.E. Kingston, D.D. Moore, J.G. Seidman, J.A. Smith, and K. Struhe. 1989. Current Protocols in Molecular Biology. Vol. 1–3. John Wiley & Sons, Inc., New York.
-
- Baksh S, Michalak M. Expression of calreticulin in Escherichia coli and identification of its Ca2+binding domains. J Biol Chem. 1991;266:21458–21465. - PubMed
-
- Bastianutto C, Clementi E, Codazzi F, Podini P, De Giorgi F, Rizzuto R, Meldolesi J, Pozzan T. Overexpression of calreticulin increases the Ca2+ capacity of rapidly exchanging Ca2+stores and reveals aspects of their lumenal microenvironment and function. J Cell Biol. 1995;130:847–855. - PMC - PubMed
-
- Brillantes AB, Ondrias K, Scott A, Kobrinsky E, Ondriasova E, Moschella MC, Jayaraman T, Lander M, Ehrlich BE, Marks AR. Stabilization of calcium release channel (ryanodine receptor) function by FK506-binding protein. Cell. 1994;77:513–523. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
