

Chlorpromazine inhibits miniature GABAergic currents by reducing the binding and by increasing the unbinding rate of GABAA receptors
- PMID: 10087062
- PMCID: PMC6786064
- DOI: 10.1523/JNEUROSCI.19-07-02474.1999
Chlorpromazine inhibits miniature GABAergic currents by reducing the binding and by increasing the unbinding rate of GABAA receptors
Abstract
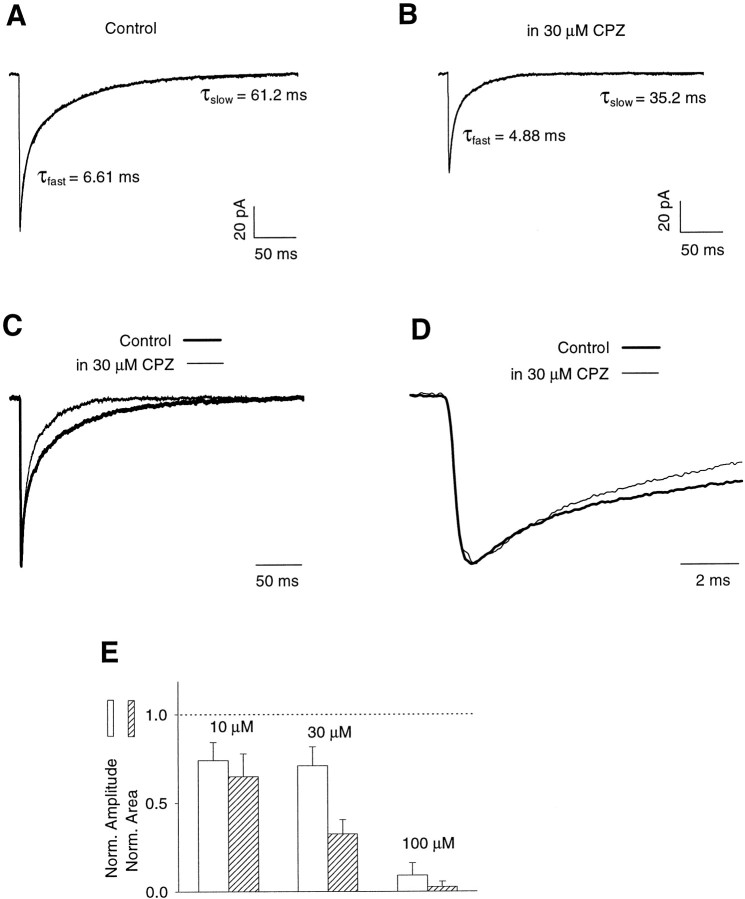
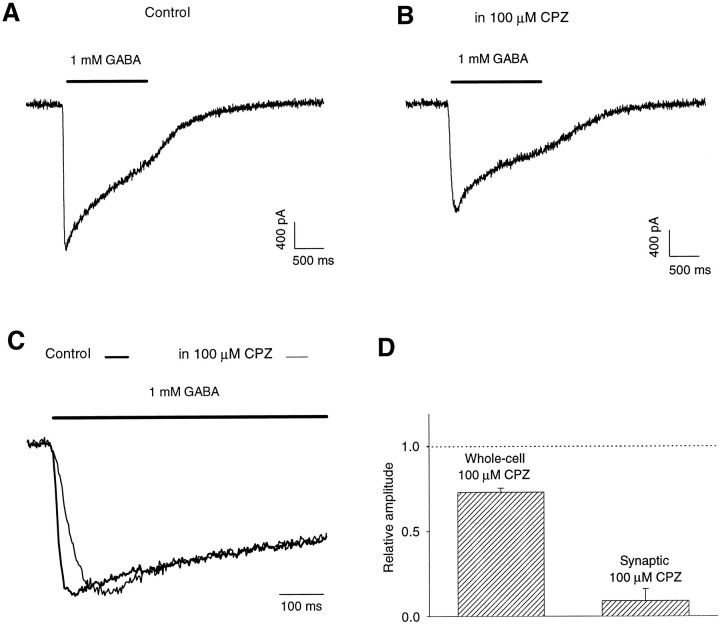
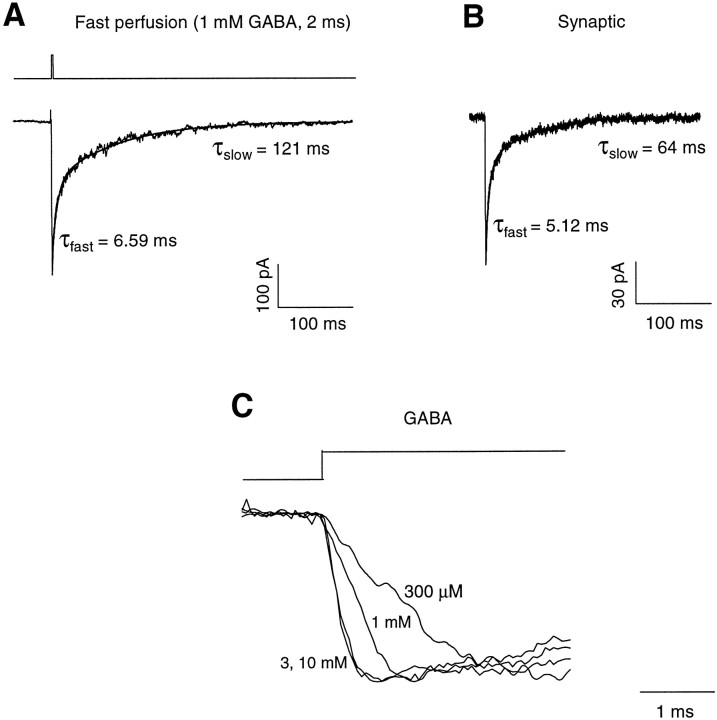
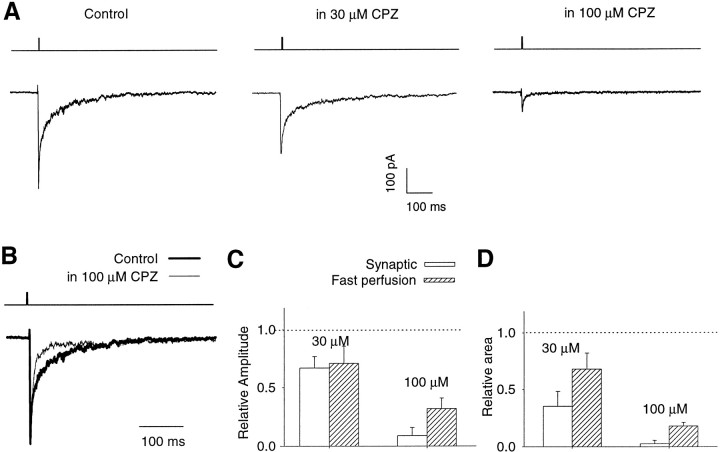
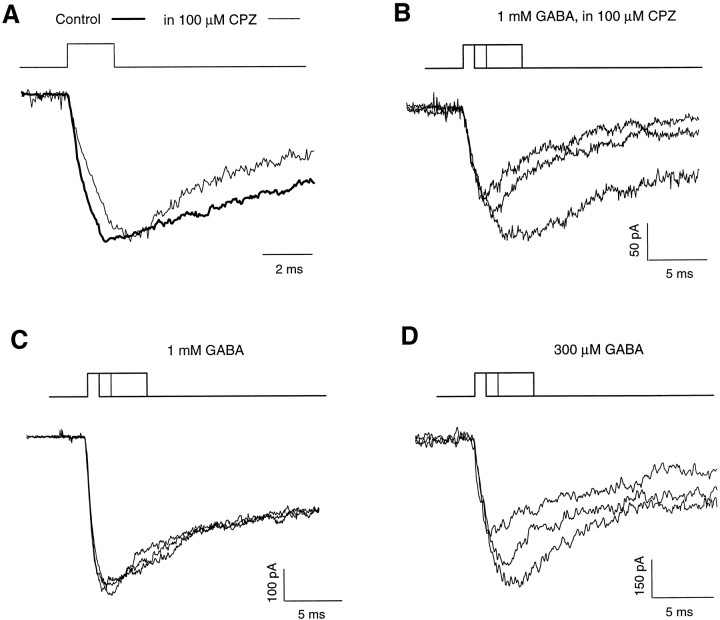
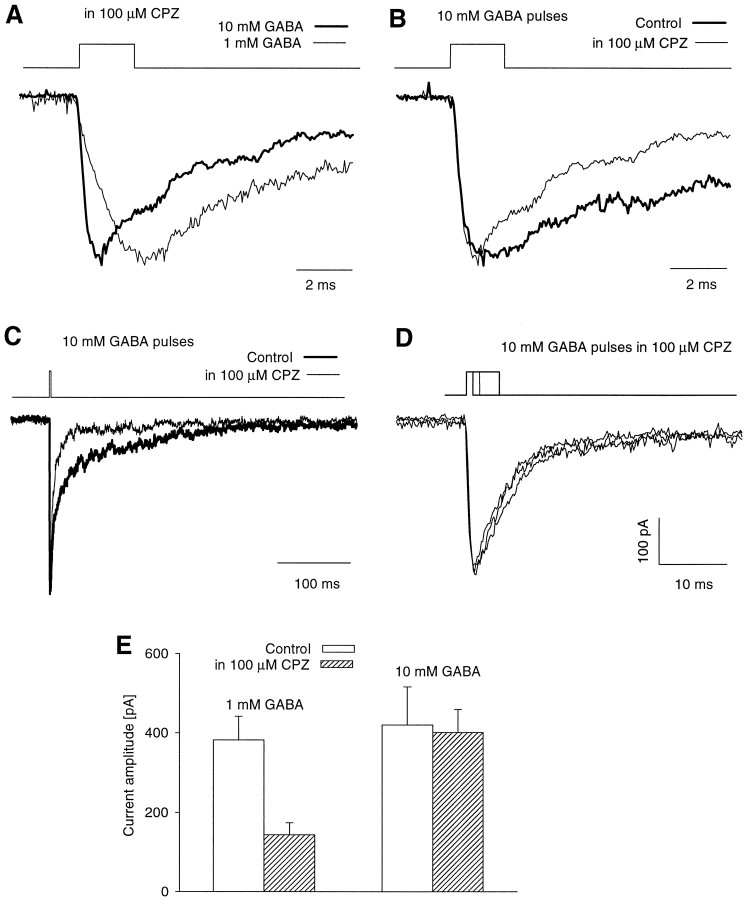
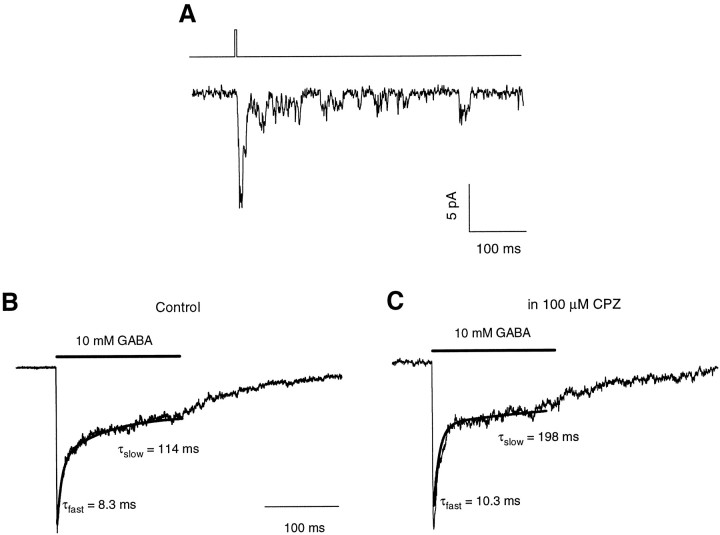
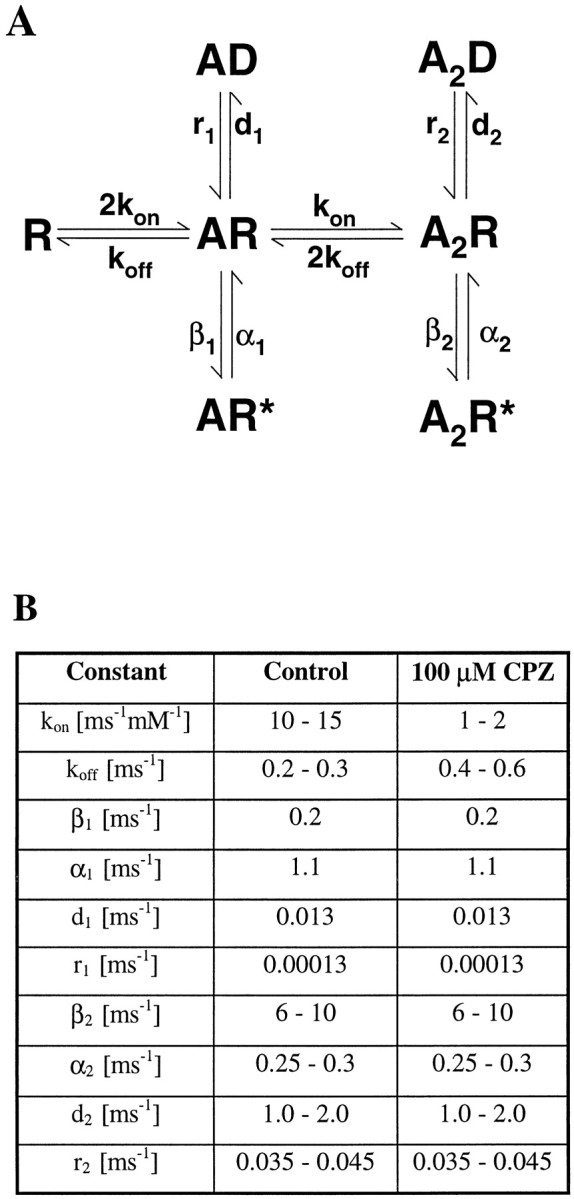
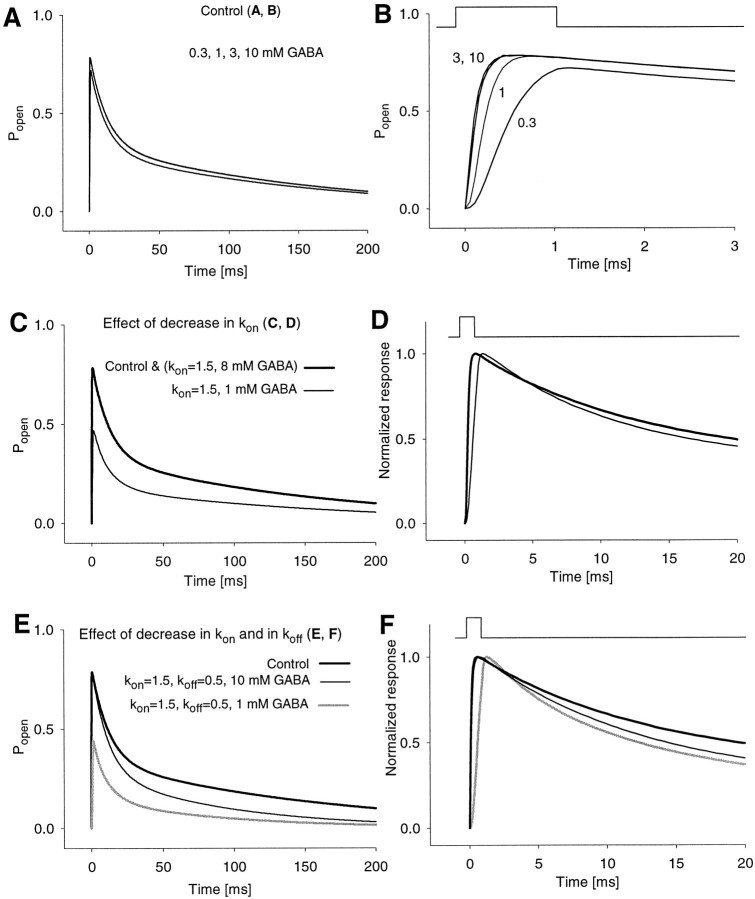
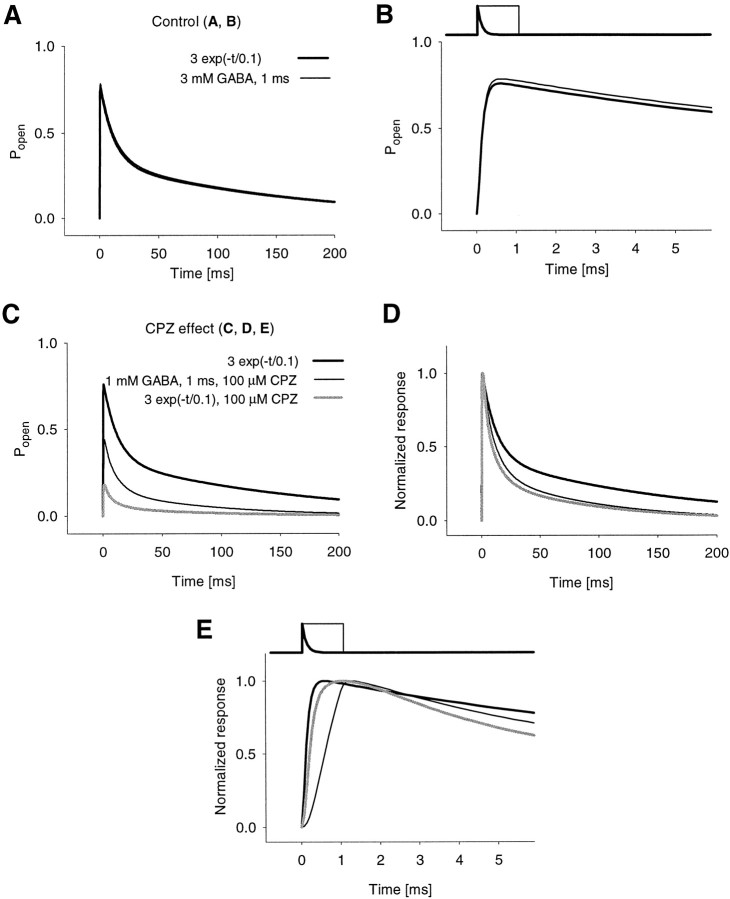
Recent studies have emphasized that nonequilibrium conditions of postsynaptic GABAA receptor (GABAAR) activation is a key factor in shaping the time course of IPSCs (Puia et al., 1994; Jones and Westbrook, 1995). Such nonequilibrium, resulting from extremely fast agonist time course, may affect the interaction between pharmacological agents and postsynaptic GABAARs. In the present study we found that chlorpromazine (CPZ), a widely used antipsychotic drug known to interfere with several ligand and voltage-gated channels, reduces the amplitude and accelerates the decay of miniature IPSCs (mIPSCs). A good qualitative reproduction of the effects of CPZ on mIPSCs was obtained when mIPSCs were mimicked by responses to ultrafast GABA applications to excised patches. Our experimental data and model simulations indicate that CPZ affects mIPSCs by decreasing the binding (kon) and by increasing the unbinding (koff) rates of GABAARs. Because of reduction of kon by CPZ, the binding reaction becomes rate-limiting, and agonist exposure of GABAARs during mIPSC is too short to activate the receptors to the same extent as in control conditions. The increase in unbinding rate is implicated as the mechanism underlying the acceleration of mIPSC decaying phase. The effect of CPZ on GABAAR binding rate, resulting in slower onset of GABA-evoked currents, provides a tool to estimate the speed of synaptic clearance of GABA. Moreover, the onset kinetics of recorded responses allowed the estimate the peak synaptic GABA concentration.
Figures
References
-
- Agopyan N, Krnjevic K. Effects of trifluoperazine on synaptically evoked potentials and membrane properties of CA1 pyramidal neurons of rat hippocampus in situ and in vitro. Synapse. 1993;13:10–19. - PubMed
-
- Benoit P, Changeux JP. Voltage dependencies of the effects of chlorpromazine on the nicotinic receptor channel from mouse muscle cell line So18. Neurosci Lett. 1993;160:81–84. - PubMed
-
- Bier M, Kits KS, Borst JGG. Relation between rise times and amplitudes of GABAergic postsynaptic currents. J Neurophysiol. 1996;75:1008–1012. - PubMed
-
- Bolotina V, Courtney KR, Khodorov B. Gate-dependent blockade of sodium channels: structure-activity relationships. Mol Pharmacol. 1992;42:423–431. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources