

Involvement of Pex13p in Pex14p localization and peroxisomal targeting signal 2-dependent protein import into peroxisomes
- PMID: 10087260
- PMCID: PMC2150583
- DOI: 10.1083/jcb.144.6.1151
Involvement of Pex13p in Pex14p localization and peroxisomal targeting signal 2-dependent protein import into peroxisomes
Abstract
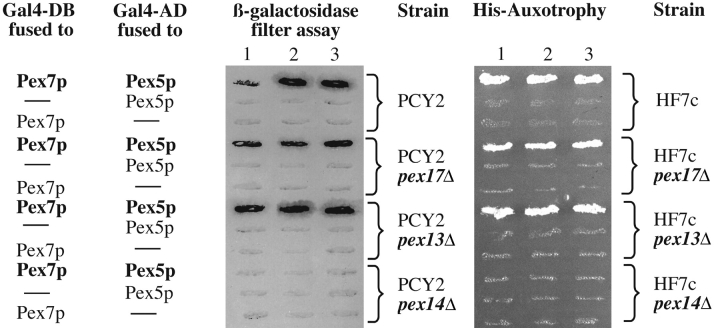
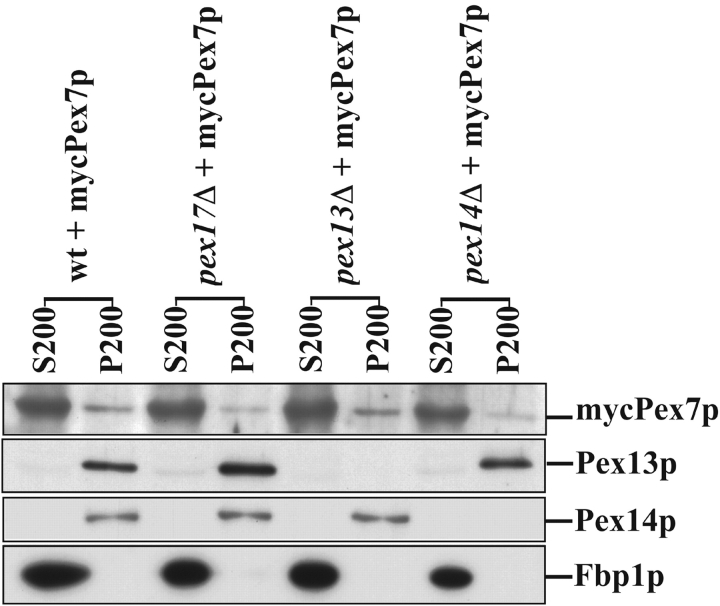
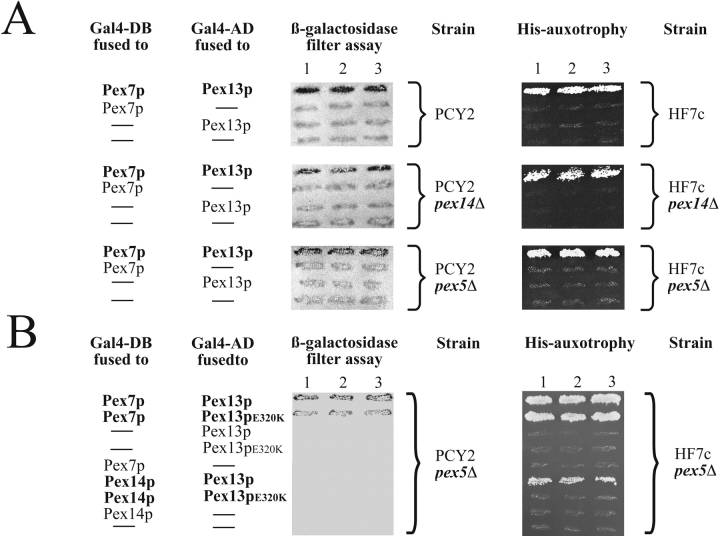
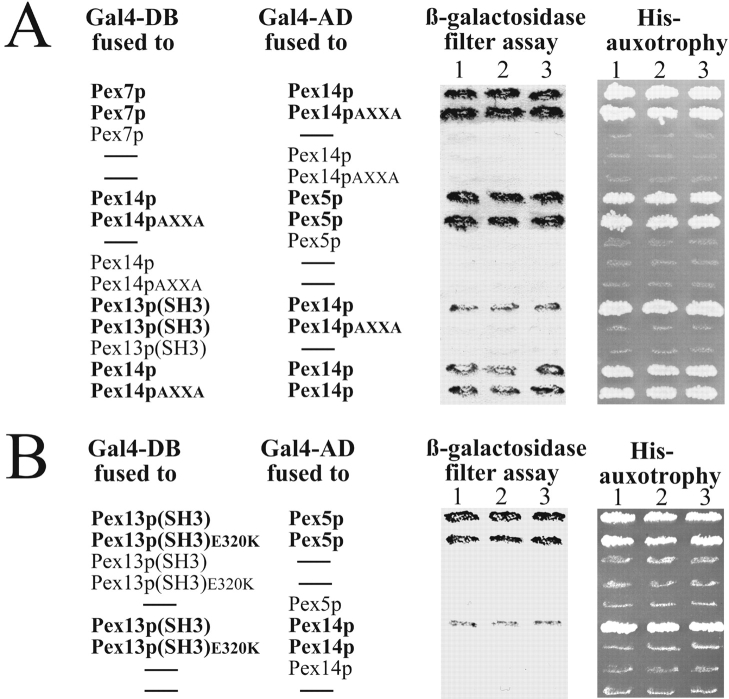
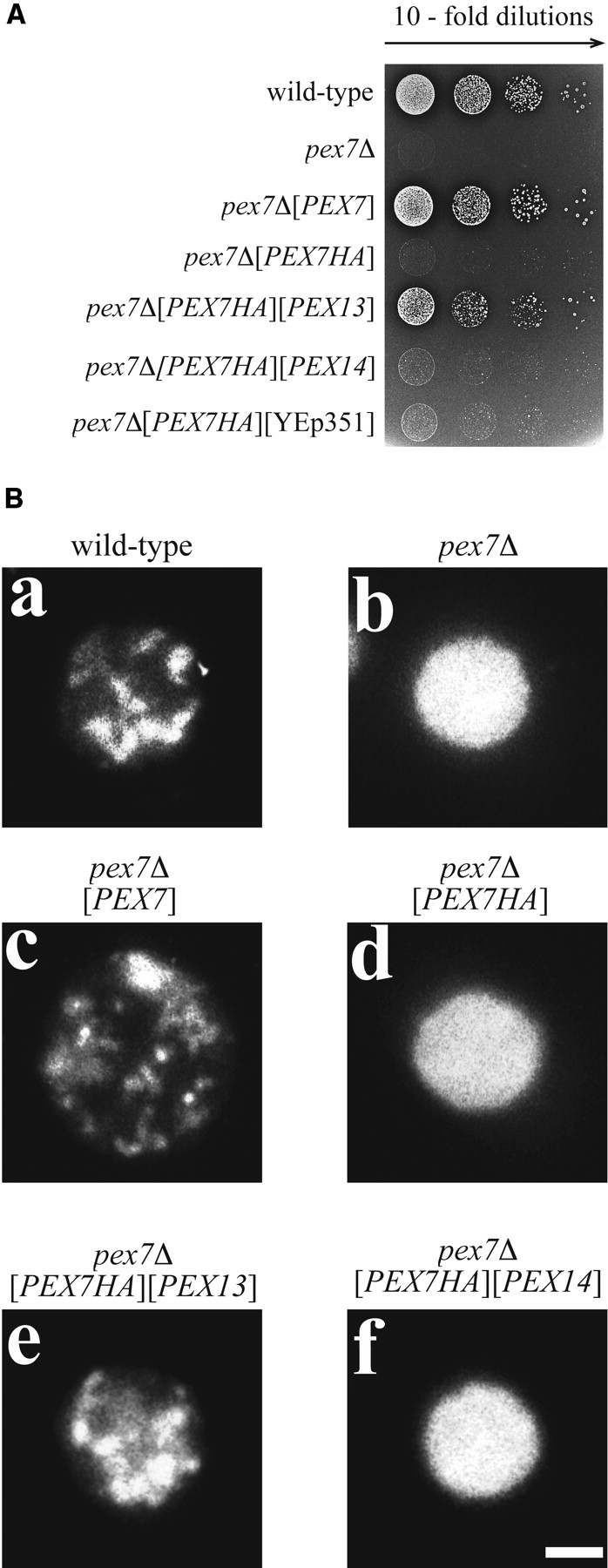
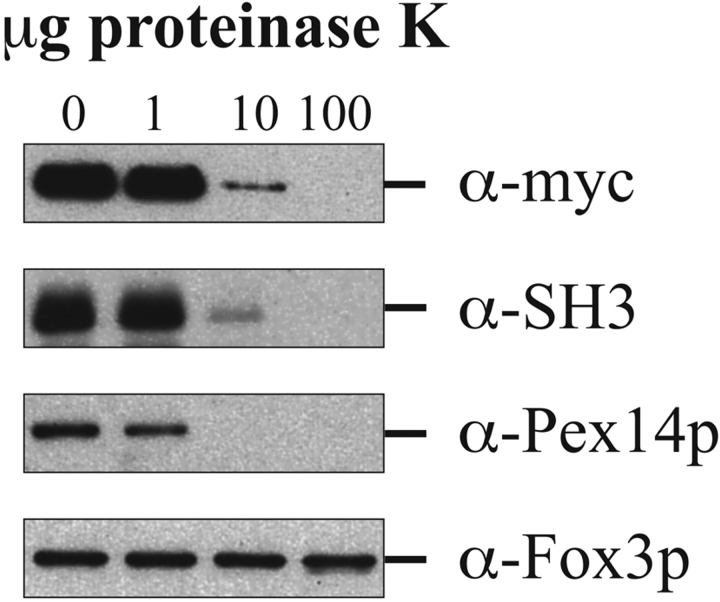
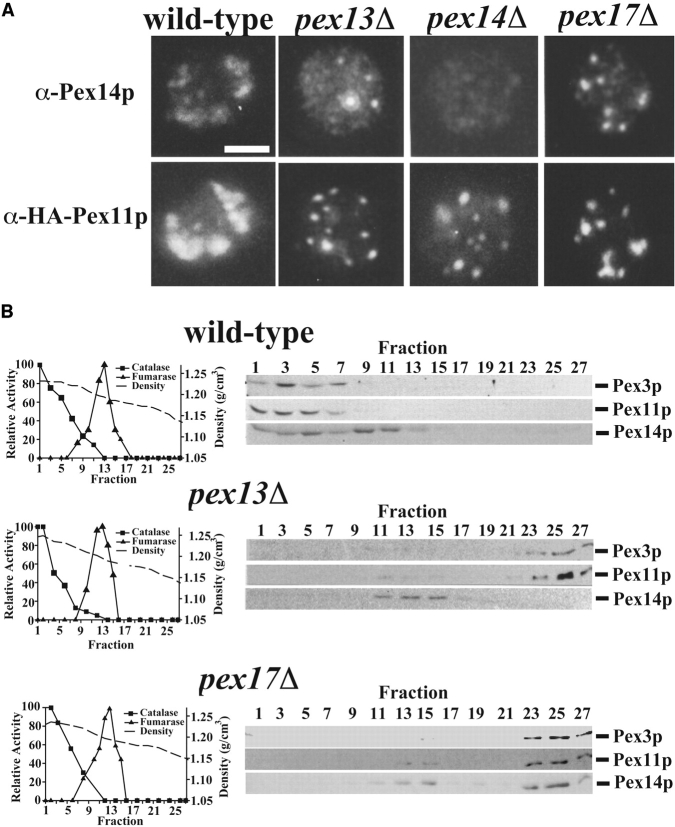
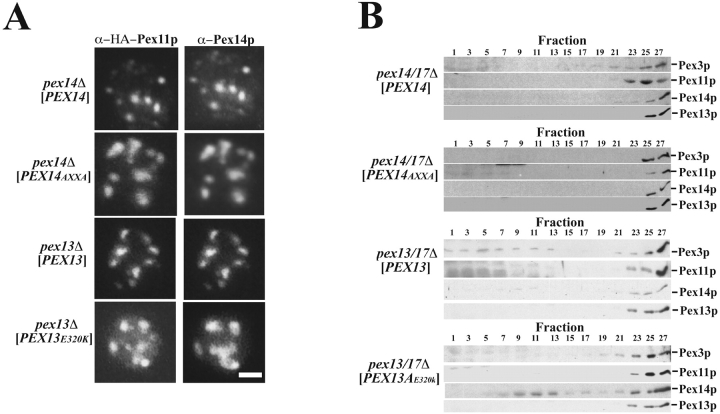
Pex13p is the putative docking protein for peroxisomal targeting signal 1 (PTS1)-dependent protein import into peroxisomes. Pex14p interacts with both the PTS1- and PTS2-receptor and may represent the point of convergence of the PTS1- and PTS2-dependent protein import pathways. We report the involvement of Pex13p in peroxisomal import of PTS2-containing proteins. Like Pex14p, Pex13p not only interacts with the PTS1-receptor Pex5p, but also with the PTS2-receptor Pex7p; however, this association may be direct or indirect. In support of distinct peroxisomal binding sites for Pex7p, the Pex7p/Pex13p and Pex7p/ Pex14p complexes can form independently. Genetic evidence for the interaction of Pex7p and Pex13p is provided by the observation that overexpression of Pex13p suppresses a loss of function mutant of Pex7p. Accordingly, we conclude that Pex7p and Pex13p functionally interact during PTS2-dependent protein import into peroxisomes. NH2-terminal regions of Pex13p are required for its interaction with the PTS2-receptor while the COOH-terminal SH3 domain alone is sufficient to mediate its interaction with the PTS1-receptor. Reinvestigation of the topology revealed both termini of Pex13p to be oriented towards the cytosol. We also found Pex13p to be required for peroxisomal association of Pex14p, yet the SH3 domain of Pex13p may not provide the only binding site for Pex14p at the peroxisomal membrane.
Figures

Similar articles
-
Peroxisomal targeting signal receptor Pex5p interacts with cargoes and import machinery components in a spatiotemporally differentiated manner: conserved Pex5p WXXXF/Y motifs are critical for matrix protein import.Mol Cell Biol. 2002 Mar;22(6):1639-55. doi: 10.1128/MCB.22.6.1639-1655.2002. Mol Cell Biol. 2002. PMID: 11865044 Free PMC article.
-
Saccharomyces cerevisiae PTS1 receptor Pex5p interacts with the SH3 domain of the peroxisomal membrane protein Pex13p in an unconventional, non-PXXP-related manner.Mol Biol Cell. 2000 Nov;11(11):3963-76. doi: 10.1091/mbc.11.11.3963. Mol Biol Cell. 2000. PMID: 11071920 Free PMC article.
-
Interactions of Pex7p and Pex18p/Pex21p with the peroxisomal docking machinery: implications for the first steps in PTS2 protein import.Mol Cell Biol. 2002 Sep;22(17):6056-69. doi: 10.1128/MCB.22.17.6056-6069.2002. Mol Cell Biol. 2002. PMID: 12167700 Free PMC article.
-
Peroxisome biogenesis and peroxisome biogenesis disorders.FEBS Lett. 2000 Jun 30;476(1-2):42-6. doi: 10.1016/s0014-5793(00)01667-7. FEBS Lett. 2000. PMID: 10878247 Review.
-
Dynamic architecture of the peroxisomal import receptor Pex5p.Biochim Biophys Acta. 2006 Dec;1763(12):1592-8. doi: 10.1016/j.bbamcr.2006.10.015. Epub 2006 Nov 10. Biochim Biophys Acta. 2006. PMID: 17141887 Review.
Cited by
-
PEX13 is mutated in complementation group 13 of the peroxisome-biogenesis disorders.Am J Hum Genet. 1999 Sep;65(3):621-34. doi: 10.1086/302534. Am J Hum Genet. 1999. PMID: 10441568 Free PMC article.
-
Pex2 and pex12 function as protein-ubiquitin ligases in peroxisomal protein import.Mol Cell Biol. 2009 Oct;29(20):5505-16. doi: 10.1128/MCB.00388-09. Epub 2009 Aug 17. Mol Cell Biol. 2009. PMID: 19687296 Free PMC article.
-
Insights into the Structure and Function of the Pex1/Pex6 AAA-ATPase in Peroxisome Homeostasis.Cells. 2022 Jun 29;11(13):2067. doi: 10.3390/cells11132067. Cells. 2022. PMID: 35805150 Free PMC article. Review.
-
The Peroxisomal PTS1-Import Defect of PEX1- Deficient Cells Is Independent of Pexophagy in Saccharomyces cerevisiae.Int J Mol Sci. 2020 Jan 29;21(3):867. doi: 10.3390/ijms21030867. Int J Mol Sci. 2020. PMID: 32013259 Free PMC article.
-
Post-translational modifications of proteins associated with yeast peroxisome membrane: An essential mode of regulatory mechanism.Genes Cells. 2021 Nov;26(11):843-860. doi: 10.1111/gtc.12892. Epub 2021 Sep 2. Genes Cells. 2021. PMID: 34472666 Free PMC article. Review.
References
-
- Albertini M, Rehling P, Erdmann R, Girzalsky W, Kiel JAKW, Veenhuis M, Kunau W-H. Pex14p, a peroxisomal membrane protein binding both receptors of the two PTS-dependent import pathways. Cell. 1997;89:83–92. - PubMed
-
- Ausubel, F.J., R. Brent, R.E. Kingston, D.D. Moore, J.G. Seidman, J.A. Smith, and K. Struhl. 1992. Current Protocols in Molecular Biology. Greene Publishing Associates, New York. 13.1.2–13.1.7.
-
- Bigl M, Escherich K. Overexpression of catalytically active yeast (Saccharomyces cerevisiae) fructose-1,6-bisphosphatase in Escherichia coli. . Biol Chem Hoppe-Seyle. 1994;375:153–160. - PubMed
-
- Blobel F, Erdmann R. Identification of a peroxisomal member of the AMP-binding protein family. Eur J Biochem. 1996;240:468–476. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
