

Extracellular matrix regulates apoptosis in mammary epithelium through a control on insulin signaling
- PMID: 10087274
- PMCID: PMC2150575
- DOI: 10.1083/jcb.144.6.1337
Extracellular matrix regulates apoptosis in mammary epithelium through a control on insulin signaling
Abstract
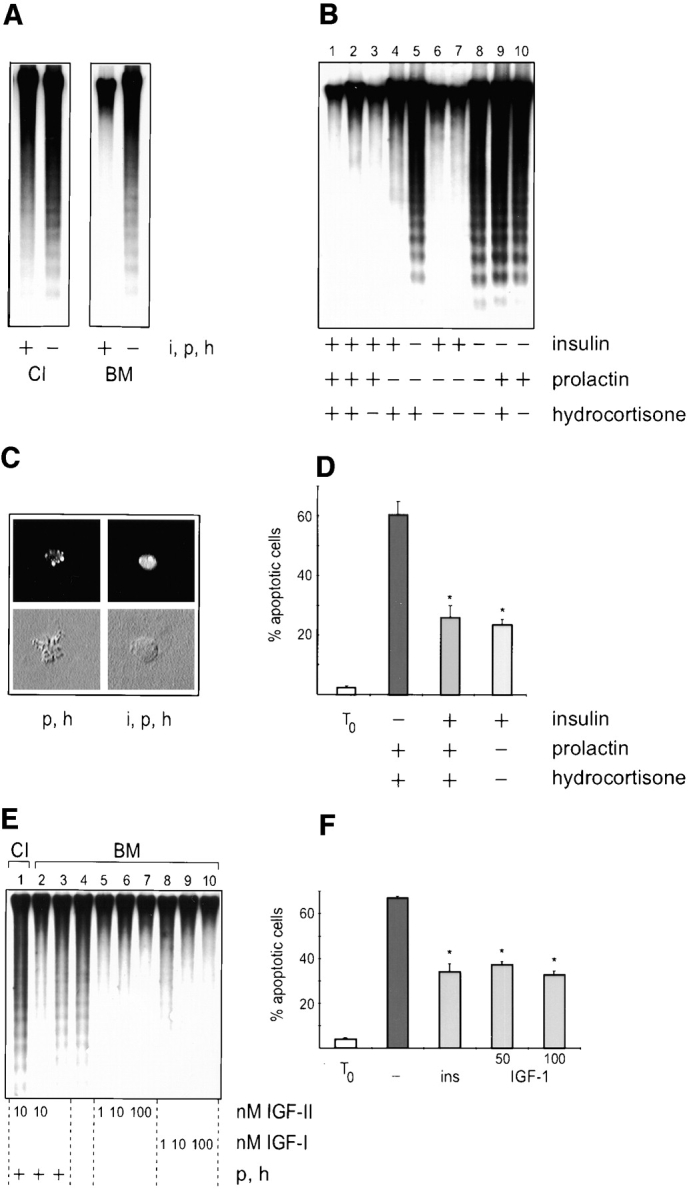
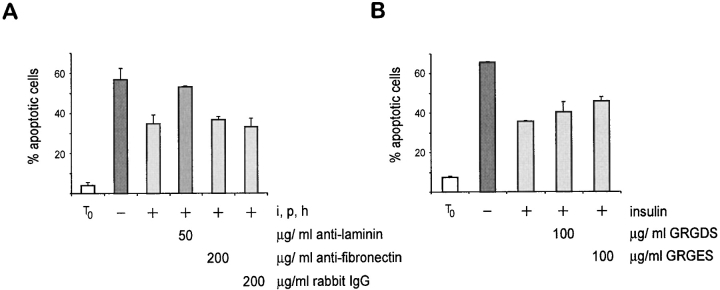
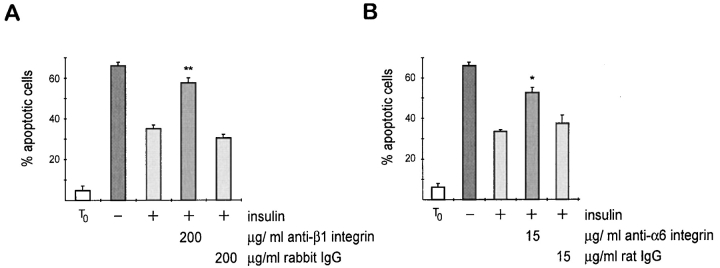
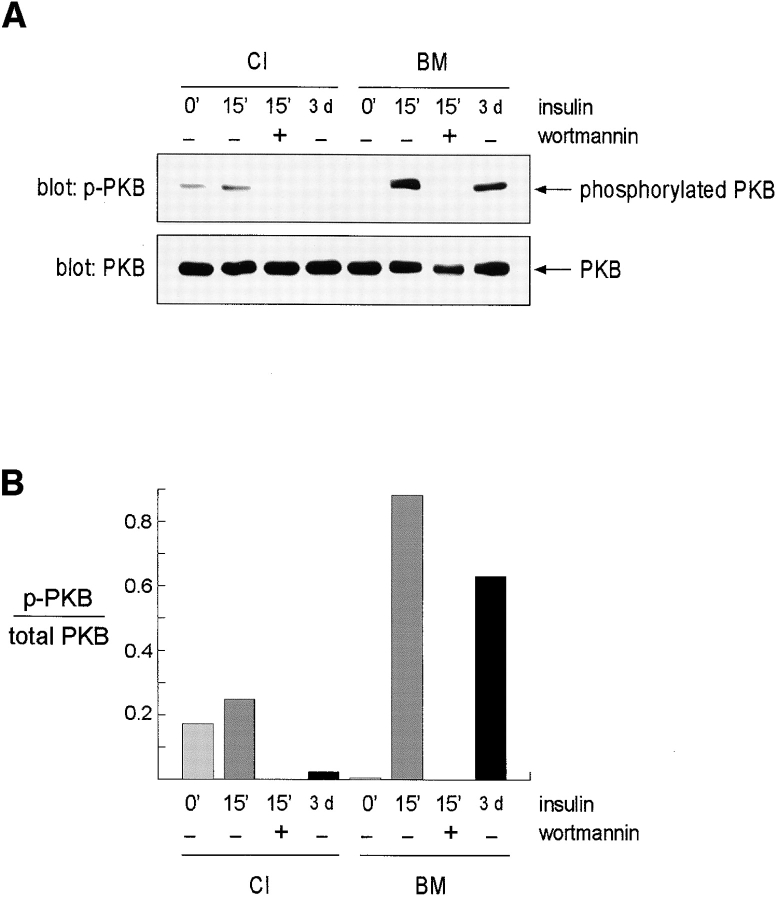
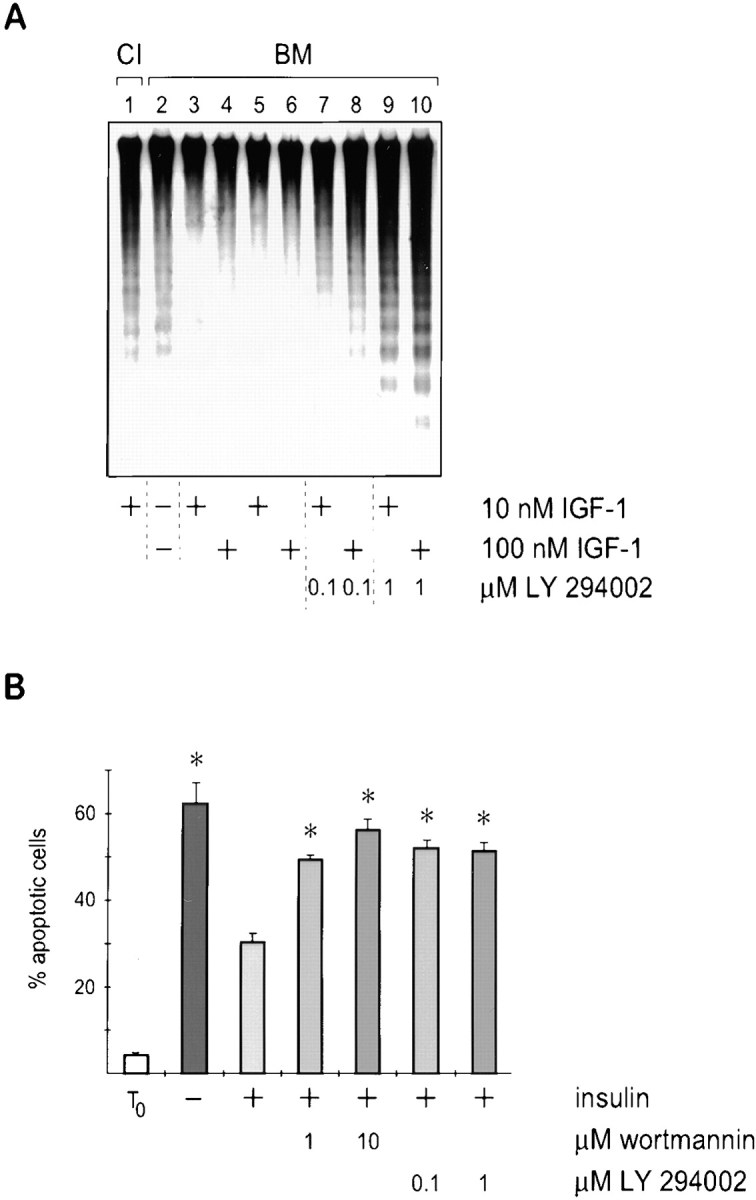
Adherent epithelial cells require interactions with the extracellular matrix for their survival, though the mechanism is ill-defined. In long term cultures of primary mammary epithelial cells, a laminin-rich basement membrane (BM) but not collagen I suppresses apoptosis, indicating that adhesion survival signals are specific in their response (. J. Cell Sci. 109:631-642). We now demonstrate that the signal from BM is mediated by integrins and requires both the alpha6 and beta1 subunits. In addition, a hormonal signal from insulin or insulin-like growth factors, but not hydrocortisone or prolactin, is necessary to suppress mammary cell apoptosis, indicating that BM and soluble factors cooperate in survival signaling. Insulin induced autophosphorylation of its receptor whether mammary cells were cultured on collagen I or BM substrata. However, both the tyrosine phosphorylation of insulin receptor substrate-1 and its association with phosphatidylinositol 3-kinase were enhanced in cells cultured on BM, as was the phosphorylation of the phosphatidylinositol 3-kinase effector, protein kinase B. These results suggest a novel extracellular matrix-dependent restriction point in insulin signaling in mammary epithelial cells. The proximal signal transduction event of insulin receptor phosphorylation is not dependent on extracellular matrix, but the activation of downstream effectors requires adhesion to BM. Since phosphatidylinositol 3-kinase was required for mammary epithelial cell survival, we propose that a possible mechanism for BM-mediated suppression of apoptosis is through its facilitative effects on insulin signaling.
Figures




References
-
- Aggeler J, Ward J, Blackie LM, Barcellos-Hoff MH, Streuli CH, Bissell MJ. Cytodifferentiation of mouse mammary epithelial cells cultured on a reconstituted basement membrane reveals striking similarities to development in vivo. J Cell Sci. 1991;99:407–417. - PubMed
-
- Bozzo C, Bellomo G, Silengo L, Tarone G, Altruda F. Soluble integrin ligands and growth factors independently rescue neuroblastoma cells from apoptosis under nonadherent conditions. Exp Cell Res. 1997;237:326–337. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
