

Initiation of peristalsis by circumferential stretch of flat sheets of guinea-pig ileum
- PMID: 10087350
- PMCID: PMC2269259
- DOI: 10.1111/j.1469-7793.1999.0525v.x
Initiation of peristalsis by circumferential stretch of flat sheets of guinea-pig ileum
Abstract
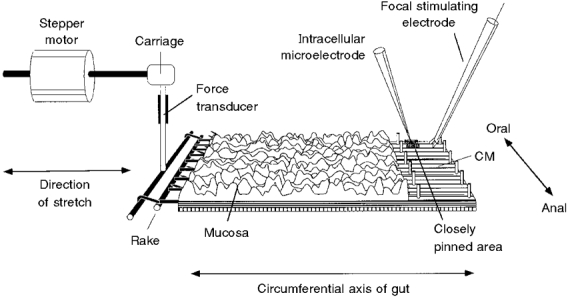
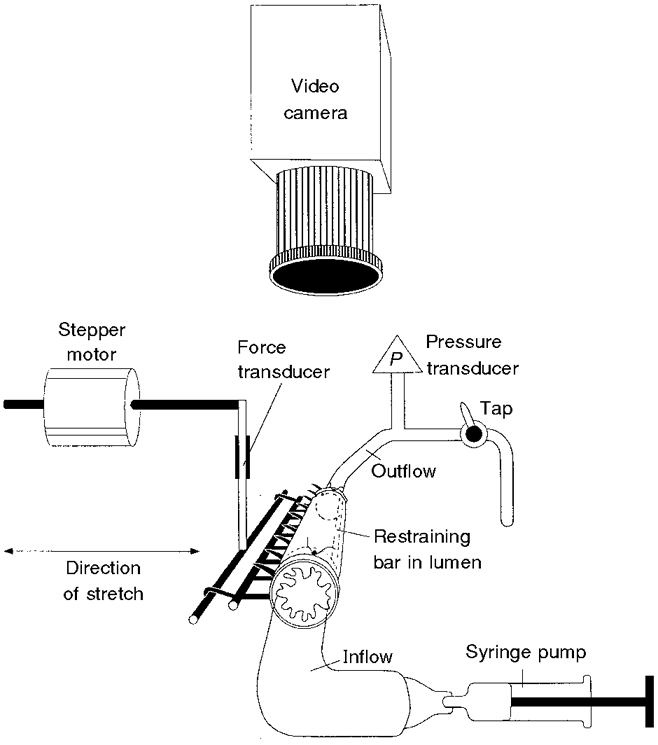
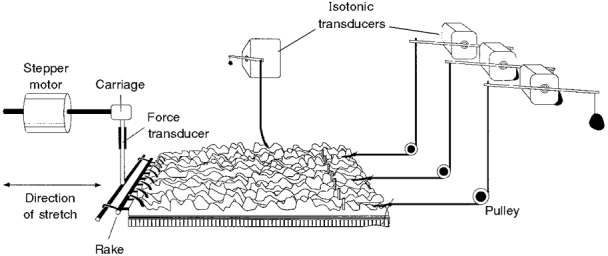
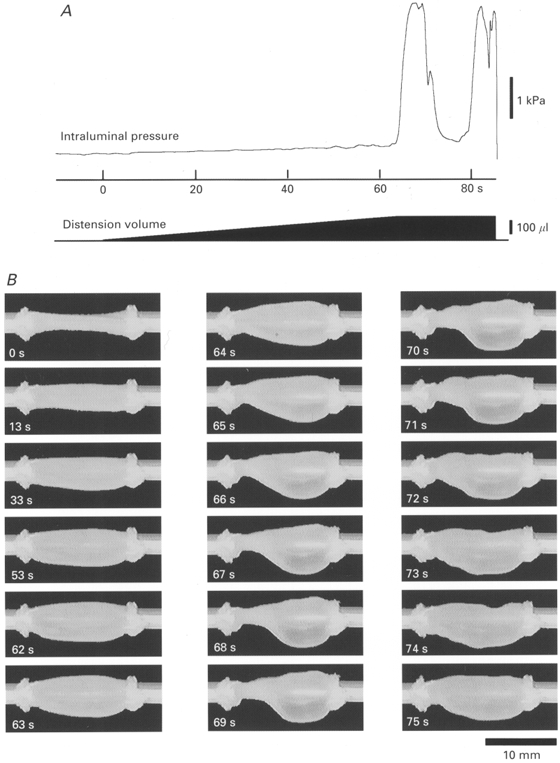
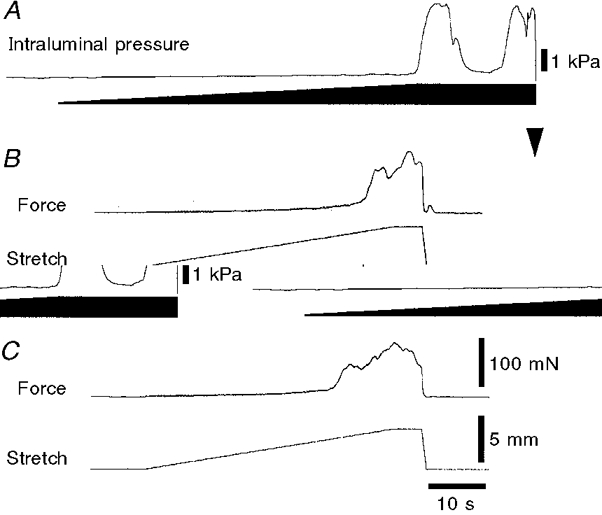
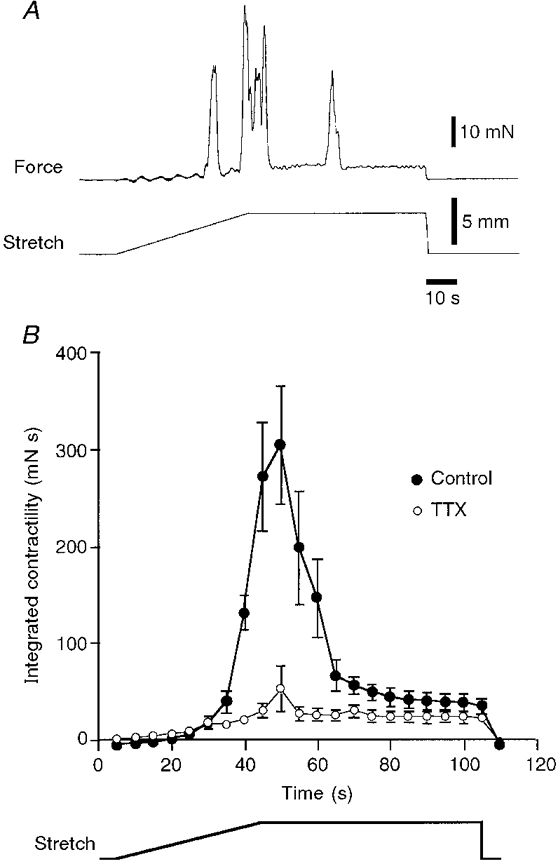
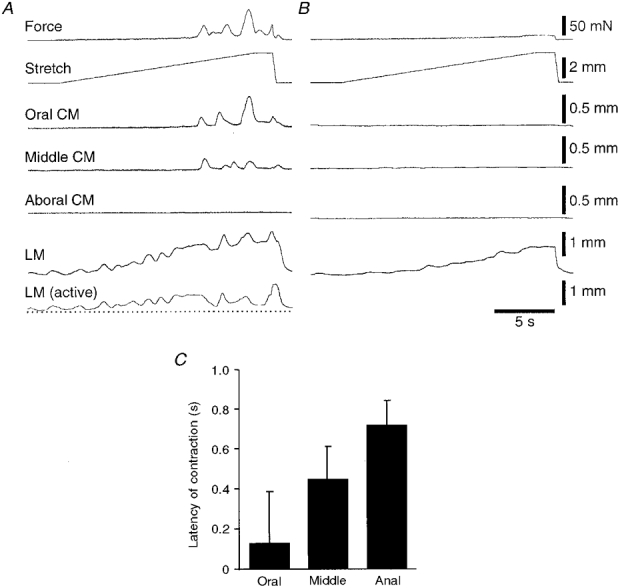
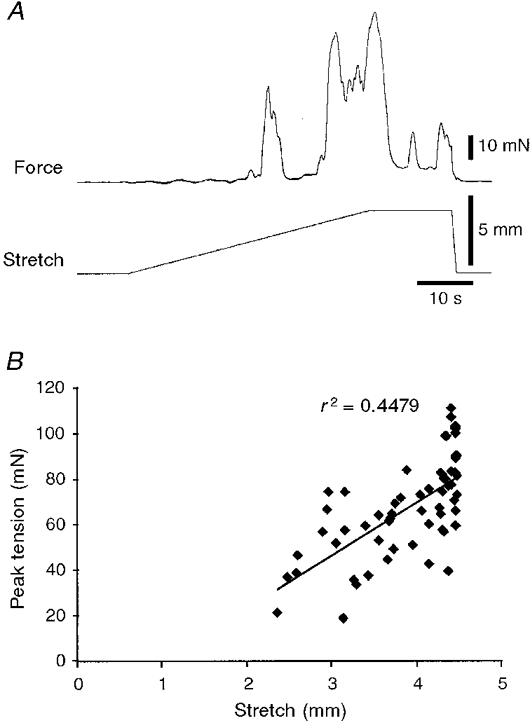
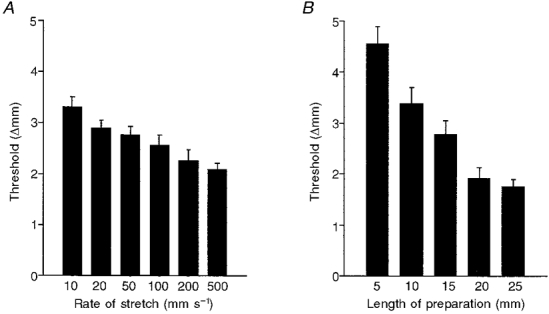
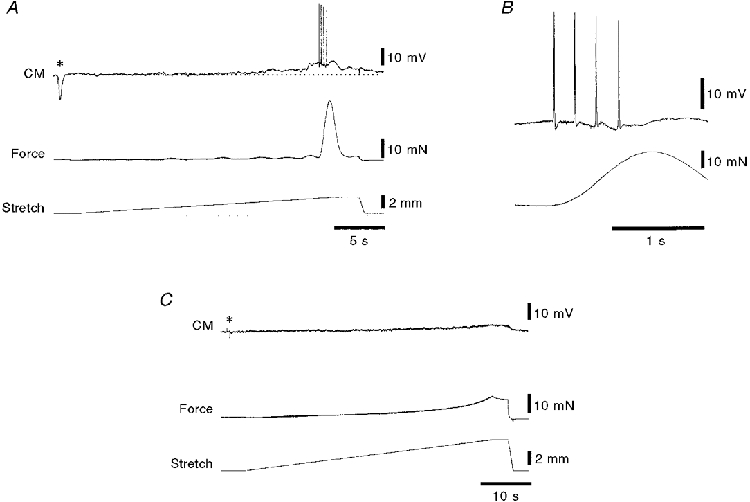
1. Segments of isolated guinea-pig intestine, 12 mm long, were distended slowly by intraluminal fluid infusion or by mechanical stretch as either a tube or flat sheet. In all cases, at a constant threshold length, a sudden, large amplitude contraction of the circular muscle occurred orally, corresponding to the initiation of peristalsis. 2. Circumferential stretch of flat sheet preparations evoked graded contractions of the longitudinal muscle (the 'preparatory phase'), which were maintained during circular muscle contraction. This suggests that the lengthening reported during the emptying phase of peristalsis is due to mechanical interactions. 3. The threshold for peristalsis was lower with more rapid stretches and was also lower in long preparations (25 mm) compared with short preparations (5-10 mm), indicating that ascending excitatory pathways play a significant role in triggering peristalsis. 4. Stretching a preparation beyond the threshold for peristalsis evoked contractions of increasing amplitude; thus peristalsis is graded above its threshold. However, during suprathreshold stretch maintained at a constant length, contractions of the circular muscle quickly declined in amplitude and frequency. 5. Circular muscle cells had a resting membrane potential approximately 6 mV more negative than the threshold for action potentials. During slow circumferential stretch, subthreshold graded excitatory motor input to the circular muscle occurred, prior to the initiation of peristalsis. However, peristalsis was initiated by a discrete large excitatory junction potential (12 +/- 2 mV) which evoked bursts of smooth muscle action potentials and which probably arose from synchronized firing of ascending excitatory neuronal pathways.
Figures
References
-
- Brookes SJH, Bywater RAR, Costa M. Characterisation of excitatory motor neurones to the circular muscle of the guinea-pig small intestine. Proceedings of the Australian Physiological and Pharmacological Society. 1995a;6:43.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources