

The role of threonine 37 in flavin reactivity of the old yellow enzyme
- PMID: 10097075
- PMCID: PMC22332
- DOI: 10.1073/pnas.96.7.3556
The role of threonine 37 in flavin reactivity of the old yellow enzyme
Abstract
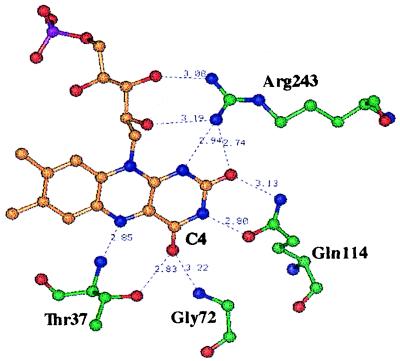
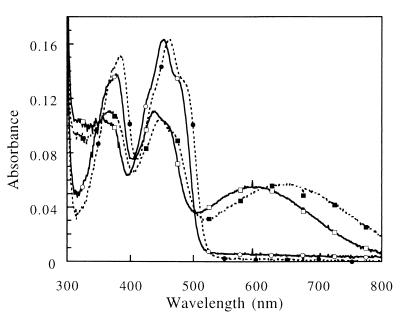
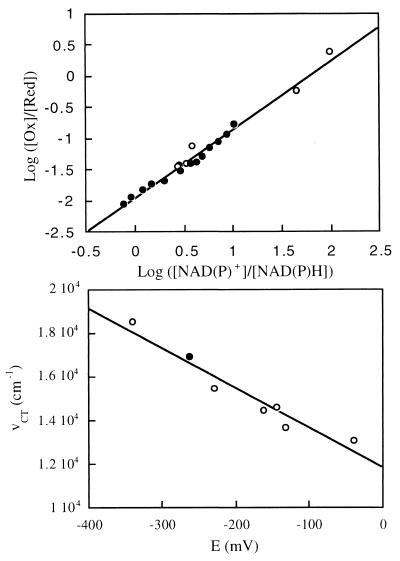
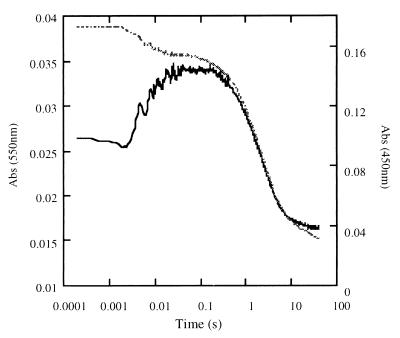
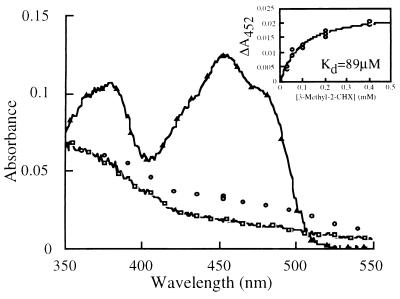
Threonine 37 is conserved among all the members of the old yellow enzyme (OYE) family. The hydroxyl group of this residue forms a hydrogen bond with the C-4 oxygen atom of the FMN reaction center of the enzyme [Fox, K. M. & Karplus, P. A. (1994) Structure 2, 1089-1105]. The position of Thr-37 and its interaction with flavin allow for speculations about its role in enzyme activity. This residue was mutated to alanine and the mutant enzyme was studied and compared with the wild-type OYE1 to evaluate its mechanistic function. The mutation has different effects on the two separate half-reactions of the enzyme. The mutant enzyme has enhanced activity in the oxidative half-reaction but the reductive half-reaction is slowed down by more than one order of magnitude. The peaks of the absorption spectra for enzyme bound with phenolic compounds are shifted toward shorter wavelengths than those of wild-type OYE1, consistent with its lower redox potential. It is suggested that Thr-37 in the wild-type OYE1 increases the redox potential of the enzyme by stabilizing the negative charge of the reduced flavin through hydrogen bonding with it.
Figures
References
-
- Warburg O, Christian W. Biochem Z. 1933;266:377–411.
-
- Theorell H. Biochem Z. 1935;275:344–346.
-
- Stott K, Saito K, Thiele D J, Massey V. J Biol Chem. 1993;268:6097–6106. - PubMed
-
- Abramovitz A S, Massey V. J Biol Chem. 1976;251:5327–5336. - PubMed
-
- Massey V, Schopfer L M. J Biol Chem. 1986;261:1215–1222. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
