

doi: 10.1073/pnas.96.7.3584.
Engineering precision RNA molecular switches
Affiliations
- PMID: 10097080
- PMCID: PMC22337
- DOI: 10.1073/pnas.96.7.3584
Item in Clipboard
Engineering precision RNA molecular switches
Proc Natl Acad Sci U S A.
.
Abstract
Ligand-specific molecular switches composed of RNA were created by coupling preexisting catalytic and receptor domains via structural bridges. Binding of ligand to the receptor triggers a conformational change within the bridge, and this structural reorganization dictates the activity of the adjoining ribozyme. The modular nature of these tripartite constructs makes possible the rapid construction of precision RNA molecular switches that trigger only in the presence of their corresponding ligand. By using similar enzyme engineering strategies, new RNA switches can be made to operate as designer molecular sensors or as a new class of genetic control elements.
Figures

Combined modular rational design and in vitro selection for FMN-sensitive allosteric ribozymes. (A) Tripartite construct consisting of a hammerhead ribozyme joined to an FMN-binding aptamer (boxed) via a random-sequence bridge composed of eight nucleotides (N). The three stems that form the unmodified ribozyme are designated I, II and III, and the site of RNA cleavage is indicated by the arrowhead. The randomized bridge serves both as a partial replacement for stem II of the ribozyme and as a flanking stem for the aptamer. The G-C base pair immediately adjacent to the catalytic core is needed for the hammerhead ribozyme to achieve maximal catalytic activity (9, 42). Selection for FMN-inducible (B) and FMN-inhibited (C) allosteric ribozymes gave rise to RNA populations that respond either positively or negatively to the presence of FMN, respectively. The initial RNA pool (G0) and successive RNA populations (G1–G6) are identified.
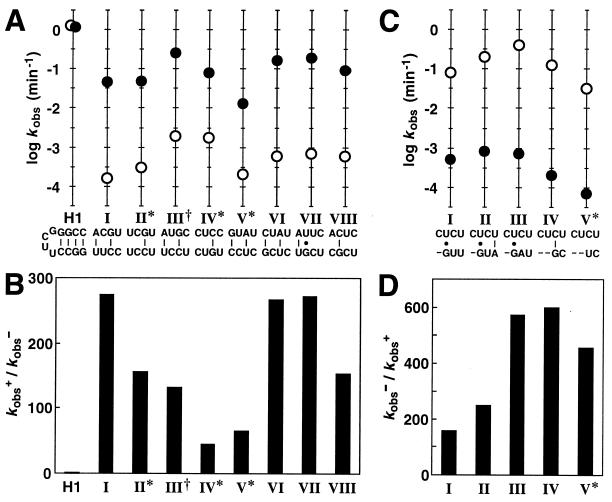
Bridge sequences and kinetic parameters for individual allosteric ribozymes. (A) Sequences and corresponding ribozyme rate constants for eight classes of induction elements isolated from G6. Plotted for each class is the logarithm of the observed rate constant for self-cleavage in the absence (○) or presence (●) of FMN. The base-pairing schemes depicted for each bridge were generated by assuming that no base-pair shift relative to the G-C base pair remaining in stem II had occurred. Indicated are classes that display greater than 20% misfolding (∗) and a class wherein an extraneous mutation exists in the stem-loop region of the aptamer domain (†). H1 is an unmodified hammerhead ribozyme (5, 8) that displays maximum catalytic activity and that remains unaffected by the presence of FMN. (B) Fold-activation of catalytic activity (kobs+/kobs−) achieved in the presence of ligand for each class of FMN-inducible ribozyme. (C) Sequences and corresponding ribozyme rate constants for five classes of inhibition elements isolated from G6. Nucleotide deletions are represented as dashes. (D) Fold-inhibition of catalytic activity (kobs−/kobs+) achieved in the presence of ligand for each class of FMN-inhibited ribozyme.
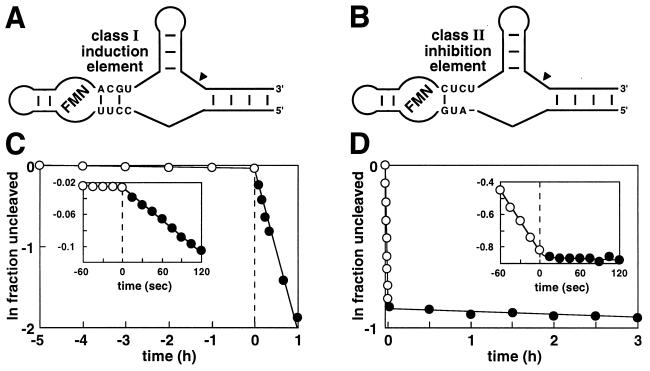
Rapid ligand-dependent modulation of allosteric ribozymes. Tripartite ribozyme constructs carrying either a class I induction element (A) or a class II inhibition element (B) are depicted. Sequences for the aptamer and ribozyme domains are as shown in Fig. 1. The performance of these ribozymes in the presence and absence of FMN are evident from plots C and D, which show the natural logarithm of the fraction of ribozyme remaining uncleaved versus time relative to FMN addition. (Insets) Plots provide an expanded view of ribozyme responses to FMN addition.

The proposed slip-structure mechanism for allosteric regulation mediated by the class I induction element (A) and class II inhibition element (B) is illustrated. Shown are the proposed stem II secondary structures of the ligand-bound and unbound states of the FMN-modulated ribozymes. Not depicted are the left- and right-flanking sequences which comprise the aptamer and ribozyme domains, respectively. Asterisks denote the G and C residues of the hammerhead ribozyme that must pair to support catalysis, and the A and G residues of the FMN aptamer that become paired on ligand binding. Also shown are bimolecular ribozyme constructs containing stem II elements designed to simulate the active or inactive slip structures proposed for the class I induction module (C; I-1 through I-3) or the class II inhibition module (D; II-1 and II-2). Thick lines identify nucleotides that form the bridge elements. Mutations made within I-3 to reinforce the desired base pairing conformation are encircled.
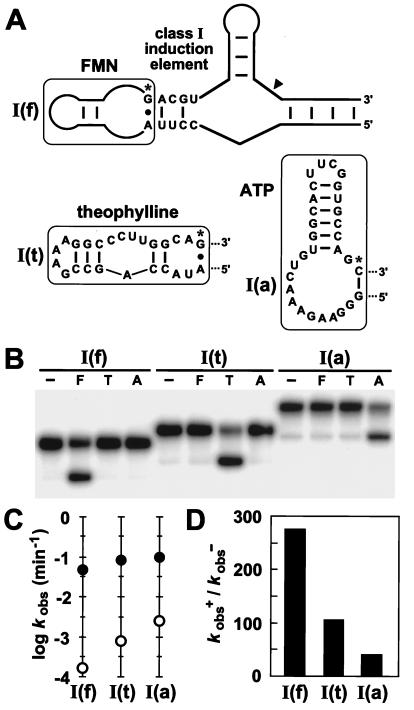
Modular characteristics of the class I induction element. (A) Sequence and secondary structures of allosteric ribozyme constructs containing either an FMN, theophylline, or ATP aptamer [constructs I(f), I(t), and I(a), respectively]. The terminal A⋅G or G-C base pairs of each aptamer (denoted by asterisks) are interactions stabilized by ligand binding. (B) Qualitative assessment of the specificity of ligand-induced ribozyme self-cleavage. Internally 32P-labeled constructs were incubated at 23°C for 15 min in the absence (−) or presence of FMN (F; 200 μM), theophylline (T; 1 mM), or ATP (A; 1 mM). (C) Kinetic parameters kobs− (○) and kobs+ (●) determined for each allosteric ribozyme construct in the absence or presence of its cognate ligand, respectively. (D) Allosteric activation of ribozyme function (kobs+/kobs−) is depicted for each construct.
References
-
- McDaniel R, Ebert-Khosla S, Hopwood D A, Khosla C. Nature (London) 1995;375:549–554. - PubMed
-
- Marsden A F A, Wilkinson B, Cortés J, Dunster N J, Staunton J, Leadlay P F. Science. 1998;279:199–202. - PubMed
-
- Hellinga H W, Marvin J S. Trends Biotechnol. 1998;16:183–189. - PubMed
-
- Crane D E, Walsh C T, Khosla C. Science. 1998;282:63–68. - PubMed
-
- Tang J, Breaker R R. Chem Biol. 1997;4:453–459. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
