

Identification of the block in targeted retroviral-mediated gene transfer
- PMID: 10097153
- PMCID: PMC22410
- DOI: 10.1073/pnas.96.7.4005
Identification of the block in targeted retroviral-mediated gene transfer
Abstract
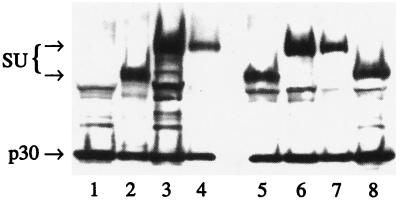
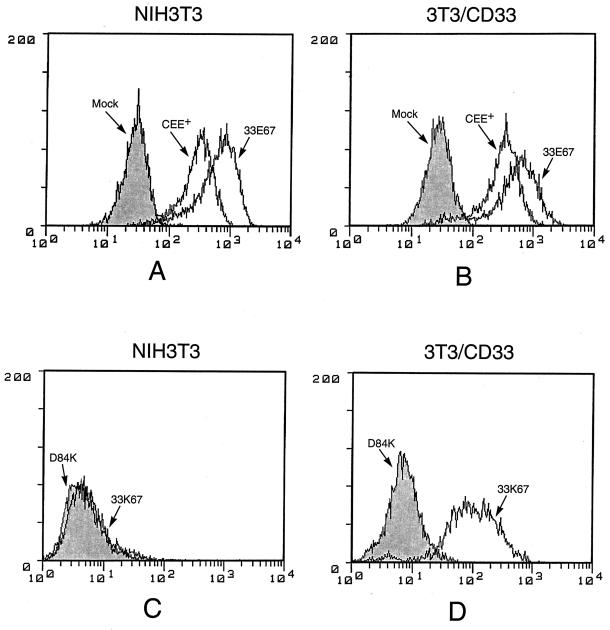
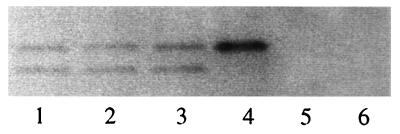
A chimeric retroviral vector (33E67) containing a CD33-specific single-chain antibody was generated in an attempt to target cells displaying the CD33 surface antigen. The chimeric envelope protein was translated, processed, and incorporated into viral particles as efficiently as wild-type envelope protein. The viral particles carrying the 33E67 envelope protein could bind efficiently to the CD33 receptor on target cells and were internalized, but no gene transfer occurred. A unique experimental approach was used to examine the basis for this postbinding block. Our data indicate that the chimeric envelope protein itself cannot participate in the fusion process, the most reasonable explanation being that this chimeric protein cannot undergo the appropriate conformational change that is thought to be triggered by receptor binding, a suggested prerequisite to subsequent fusion and core entry. These results indicate that the block to gene transfer in this system, and probably in most of the current chimeric retroviral vectors to date, is the inability of the chimeric envelope protein to undergo this obligatory conformational change.
Figures




Similar articles
-
Retroviral vector-mediated gene transfer into umbilical cord blood CD34brCD38-CD33- cells.Exp Hematol. 1999 Aug;27(8):1244-54. doi: 10.1016/s0301-472x(99)00065-x. Exp Hematol. 1999. PMID: 10428501
-
Correction of cross-linker sensitivity of Fanconi anemia group F cells by CD33-mediated protein transfer.Blood. 2001 Dec 15;98(13):3817-22. doi: 10.1182/blood.v98.13.3817. Blood. 2001. PMID: 11739191
-
Gamma-retroviral vectors enveloped with an antibody and an engineered fusogenic protein achieved antigen-specific targeting.Biotechnol Bioeng. 2008 Oct 1;101(2):357-68. doi: 10.1002/bit.21903. Biotechnol Bioeng. 2008. PMID: 18435481 Free PMC article.
-
Modified envelope glycoproteins to retarget retroviral vectors.Curr Gene Ther. 2003 Oct;3(5):405-10. doi: 10.2174/1566523034578267. Curr Gene Ther. 2003. PMID: 14529348 Review.
-
Chimeric T-cell receptors: new challenges for targeted immunotherapy in hematologic malignancies.Haematologica. 2007 Mar;92(3):381-8. doi: 10.3324/haematol.10873. Haematologica. 2007. PMID: 17339188 Review.
Cited by
-
Two point mutations produce infectious retrovirus bearing a green fluorescent protein-SU fusion protein.J Virol. 2001 Dec;75(23):11881-5. doi: 10.1128/JVI.75.23.11881-11885.2001. J Virol. 2001. PMID: 11689670 Free PMC article.
-
Versatile targeting system for lentiviral vectors involving biotinylated targeting molecules.Virology. 2018 Dec;525:170-181. doi: 10.1016/j.virol.2018.09.017. Epub 2018 Oct 2. Virology. 2018. PMID: 30290312 Free PMC article.
-
Single-chain antibody displayed on a recombinant measles virus confers entry through the tumor-associated carcinoembryonic antigen.J Virol. 2001 Mar;75(5):2087-96. doi: 10.1128/JVI.75.5.2087-2096.2001. J Virol. 2001. PMID: 11160713 Free PMC article.
-
Functional characterization of the N termini of murine leukemia virus envelope proteins.J Virol. 2001 May;75(9):4357-66. doi: 10.1128/JVI.75.9.4357-4366.2001. J Virol. 2001. PMID: 11287584 Free PMC article.
-
Receptors and entry cofactors for retroviruses include single and multiple transmembrane-spanning proteins as well as newly described glycophosphatidylinositol-anchored and secreted proteins.Microbiol Mol Biol Rev. 2001 Sep;65(3):371-89, table of contents. doi: 10.1128/MMBR.65.3.371-389.2001. Microbiol Mol Biol Rev. 2001. PMID: 11528001 Free PMC article. Review.
References
-
- Anderson W F. Nature (London) 1998;392:25–30. - PubMed
-
- Ager S, Nilson B H K, Morling F J, Peng K W, Cosset F-L, Russell S J. Hum Gene Ther. 1996;7:2157–2164. - PubMed
-
- Goud B, Legrain P, Buttin G. Virology. 1988;163:251–254. - PubMed
-
- Kasahara N, Dozy A M, Kan Y W. Science. 1994;266:1373–1376. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
