

Modulation of nicotinic acetylcholine receptors by strychnine
- PMID: 10097172
- PMCID: PMC22429
- DOI: 10.1073/pnas.96.7.4113
Modulation of nicotinic acetylcholine receptors by strychnine
Abstract
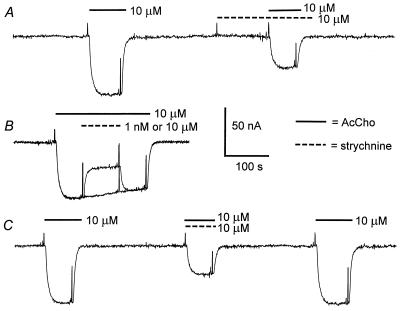
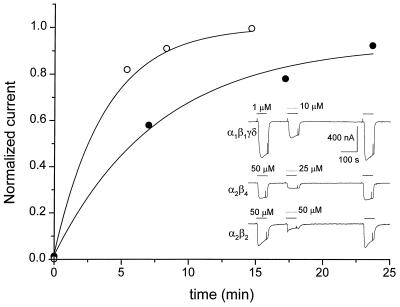
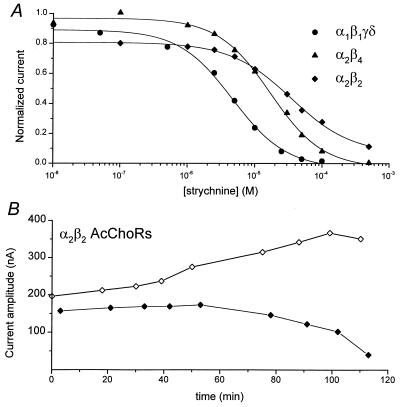
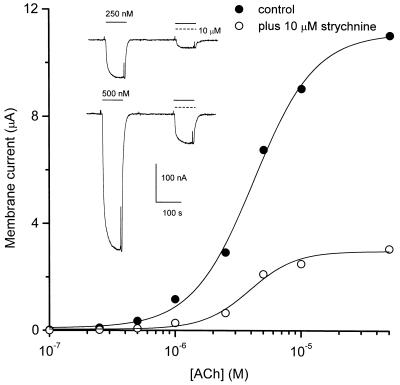
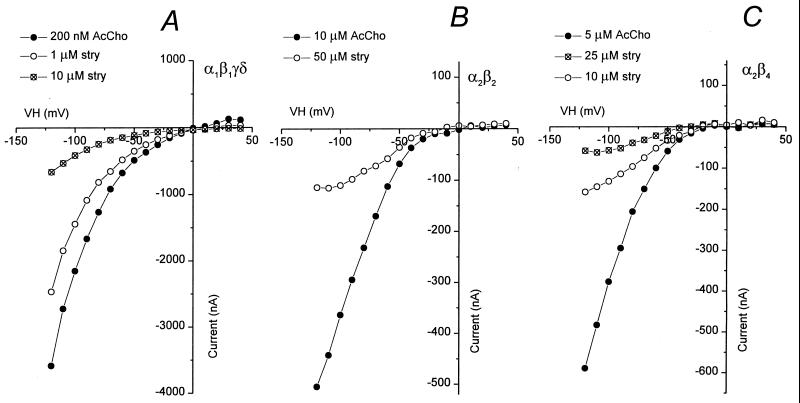
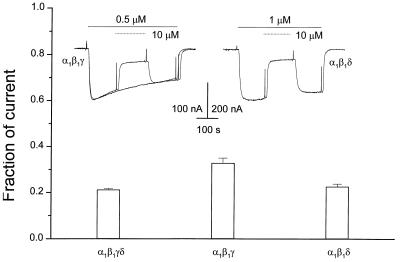
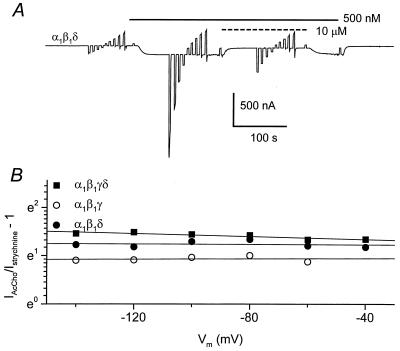
Strychnine, a potent and selective antagonist at glycine receptors, was found to inhibit muscle (alpha1beta1gammadelta, alpha1beta1gamma, and alpha1beta1delta) and neuronal (alpha2beta2 and alpha2beta4) nicotinic acetylcholine receptors (AcChoRs) expressed in Xenopus oocytes. Strychnine alone (up to 500 microM) did not elicit membrane currents in oocytes expressing AcChoRs, but, when applied before, concomitantly, or during superfusion of acetylcholine (AcCho), it rapidly and reversibly inhibited the current elicited by AcCho (AcCho-current). Although in the three cases the AcCho-current was reduced to the same level, its recovery was slower when the oocytes were preincubated with strychnine. The amount of AcCho-current inhibition depended on the receptor subtype, and the order of blocking potency by strychnine was alpha1beta1gammadelta > alpha2beta4 > alpha2beta2. With the three forms of drug application, the Hill coefficient was close to one, suggesting a single site for the receptor interaction with strychnine, and this interaction appears to be noncompetitive. The inhibitory effects on muscle AcChoRs were voltage-independent, and the apparent dissociation constant for AcCho was not appreciably changed by strychnine. In contrast, the inhibitory effects on neuronal AcChoRs were voltage-dependent, with an electrical distance of approximately 0.35. We conclude that strychnine regulates reversibly and noncompetitively the embryonic type of muscle AcChoR and some forms of neuronal AcChoRs. In the former case, strychnine presumably inhibits allosterically the receptor by binding at an external domain whereas, in the latter case, it blocks the open receptor-channel complex.
Figures
Similar articles
-
Antagonism of nicotinic acetylcholine receptors by inhibitors of monoamine uptake.Mol Psychiatry. 2001 Sep;6(5):511-9. doi: 10.1038/sj.mp.4000885. Mol Psychiatry. 2001. PMID: 11526465
-
Activation and blocking of neuronal nicotinic acetylcholine receptor reconstituted in Xenopus oocytes.Proc Natl Acad Sci U S A. 1990 Mar;87(5):1993-7. doi: 10.1073/pnas.87.5.1993. Proc Natl Acad Sci U S A. 1990. PMID: 1968642 Free PMC article.
-
Strychnine activates neuronal alpha7 nicotinic receptors after mutations in the leucine ring and transmitter binding site domains.Proc Natl Acad Sci U S A. 1999 Nov 9;96(23):13421-6. doi: 10.1073/pnas.96.23.13421. Proc Natl Acad Sci U S A. 1999. PMID: 10557336 Free PMC article.
-
Inhibition of neuronal nicotinic acetylcholine receptors by La(3+).Eur J Pharmacol. 2002 Apr 19;441(1-2):15-21. doi: 10.1016/s0014-2999(01)01522-9. Eur J Pharmacol. 2002. PMID: 12007916
-
Strychnine and its mono- and dimeric analogues: a pharmaco-chemical perspective.Nat Prod Rep. 2022 Oct 19;39(10):1910-1937. doi: 10.1039/d1np00079a. Nat Prod Rep. 2022. PMID: 35380133 Review.
Cited by
-
Role of glycine in nociceptive and non-nociceptive bladder reflexes and pudendal afferent inhibition of these reflexes in cats.Neurourol Urodyn. 2016 Sep;35(7):798-804. doi: 10.1002/nau.22821. Epub 2015 Jul 5. Neurourol Urodyn. 2016. PMID: 26147494 Free PMC article.
-
Engineering a surrogate human heteromeric α/β glycine receptor orthosteric site exploiting the structural homology and stability of acetylcholine-binding protein.IUCrJ. 2019 Sep 4;6(Pt 6):1014-1023. doi: 10.1107/S205225251901114X. eCollection 2019 Nov 1. IUCrJ. 2019. PMID: 31709057 Free PMC article.
-
Fast synaptic transmission mediated by alpha-bungarotoxin-sensitive nicotinic acetylcholine receptors in lamina X neurones of neonatal rat spinal cord.J Physiol. 2002 Nov 1;544(3):727-39. doi: 10.1113/jphysiol.2002.028894. J Physiol. 2002. PMID: 12411519 Free PMC article.
-
VirtuousPocketome: a computational tool for screening protein-ligand complexes to identify similar binding sites.Sci Rep. 2024 Mar 15;14(1):6296. doi: 10.1038/s41598-024-56893-7. Sci Rep. 2024. PMID: 38491261 Free PMC article.
-
Exploring neurotransmitters and their receptors for breast cancer prevention and treatment.Theranostics. 2023 Jan 31;13(3):1109-1129. doi: 10.7150/thno.81403. eCollection 2023. Theranostics. 2023. PMID: 36793869 Free PMC article. Review.
References
-
- Cockcroft V B, Ortels M, Lunt G G. Ann N Y Acad Sci. 1995;757:40–47. - PubMed
-
- Sargent P B. Annu Rev Neurosci. 1993;16:403–443. - PubMed
-
- McGehee D S, Role L W. Annu Rev Physiol. 1995;57:521–546. - PubMed
-
- Karlin A, Akabas M H. Neuron. 1995;15:1231–1244. - PubMed
-
- Galzi J L, Revah F, Bessis A, Changeux J P. Annu Rev Pharmacol Toxicol. 1991;31:37–72. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
