

Cis-regulatory elements of the mitotic regulator, string/Cdc25
- PMID: 10101114
- PMCID: PMC10176497
- DOI: 10.1242/dev.126.9.1793
Cis-regulatory elements of the mitotic regulator, string/Cdc25
Abstract
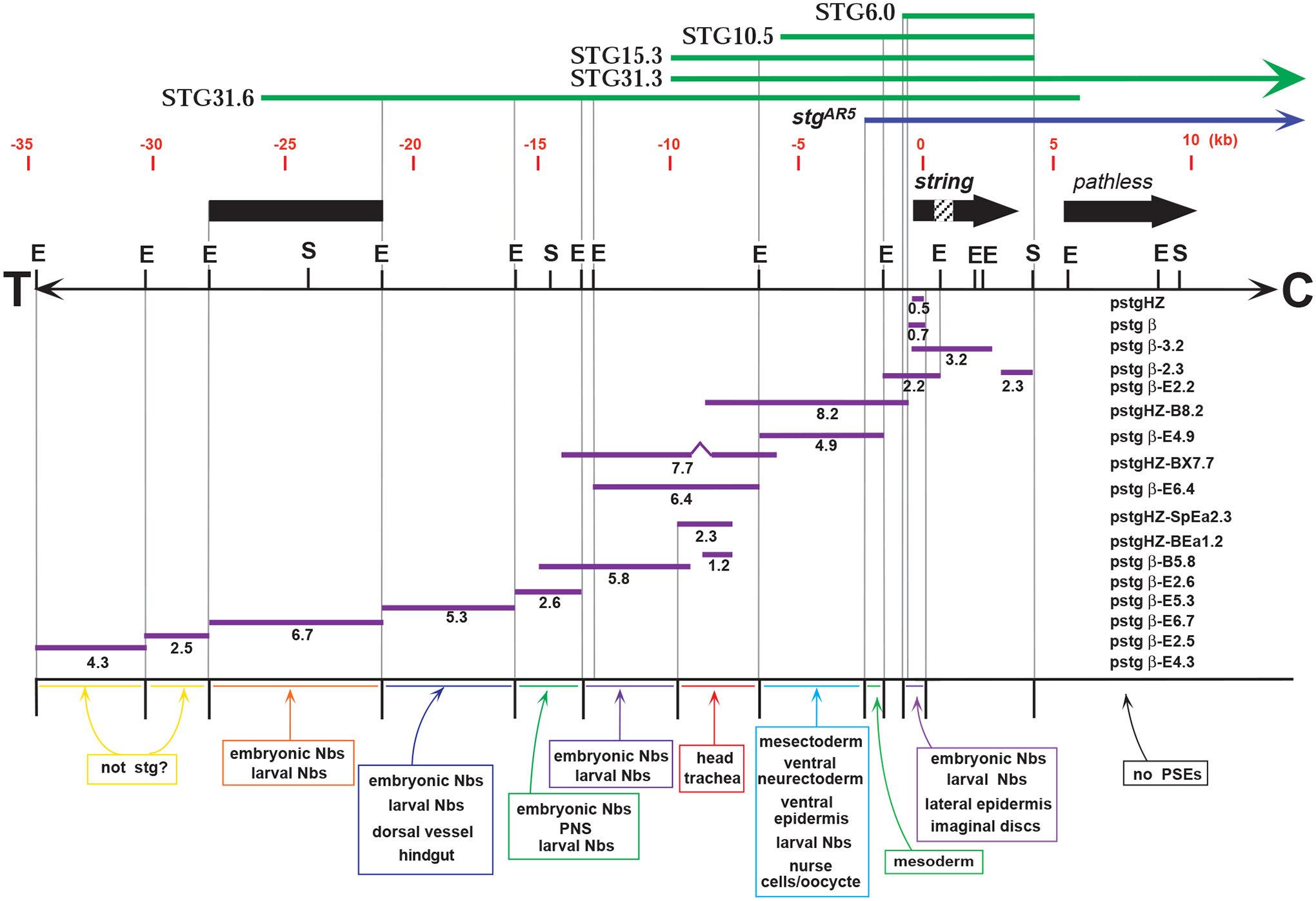
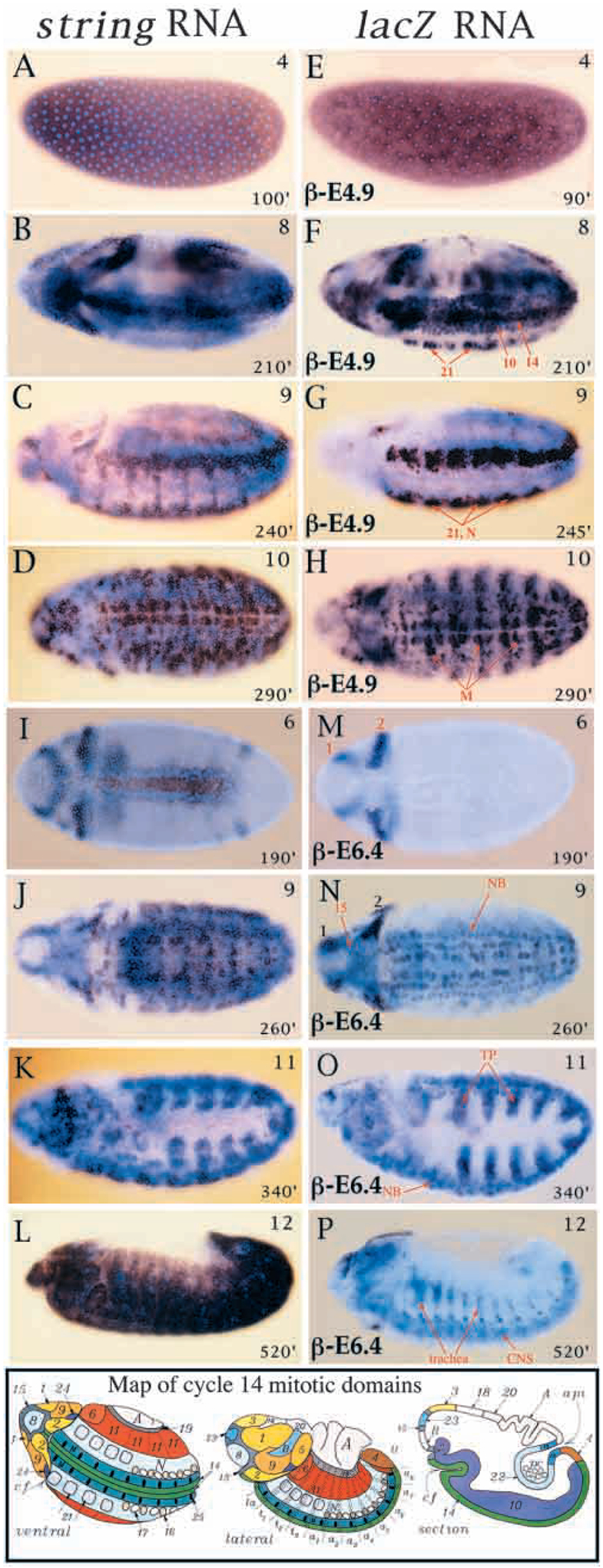
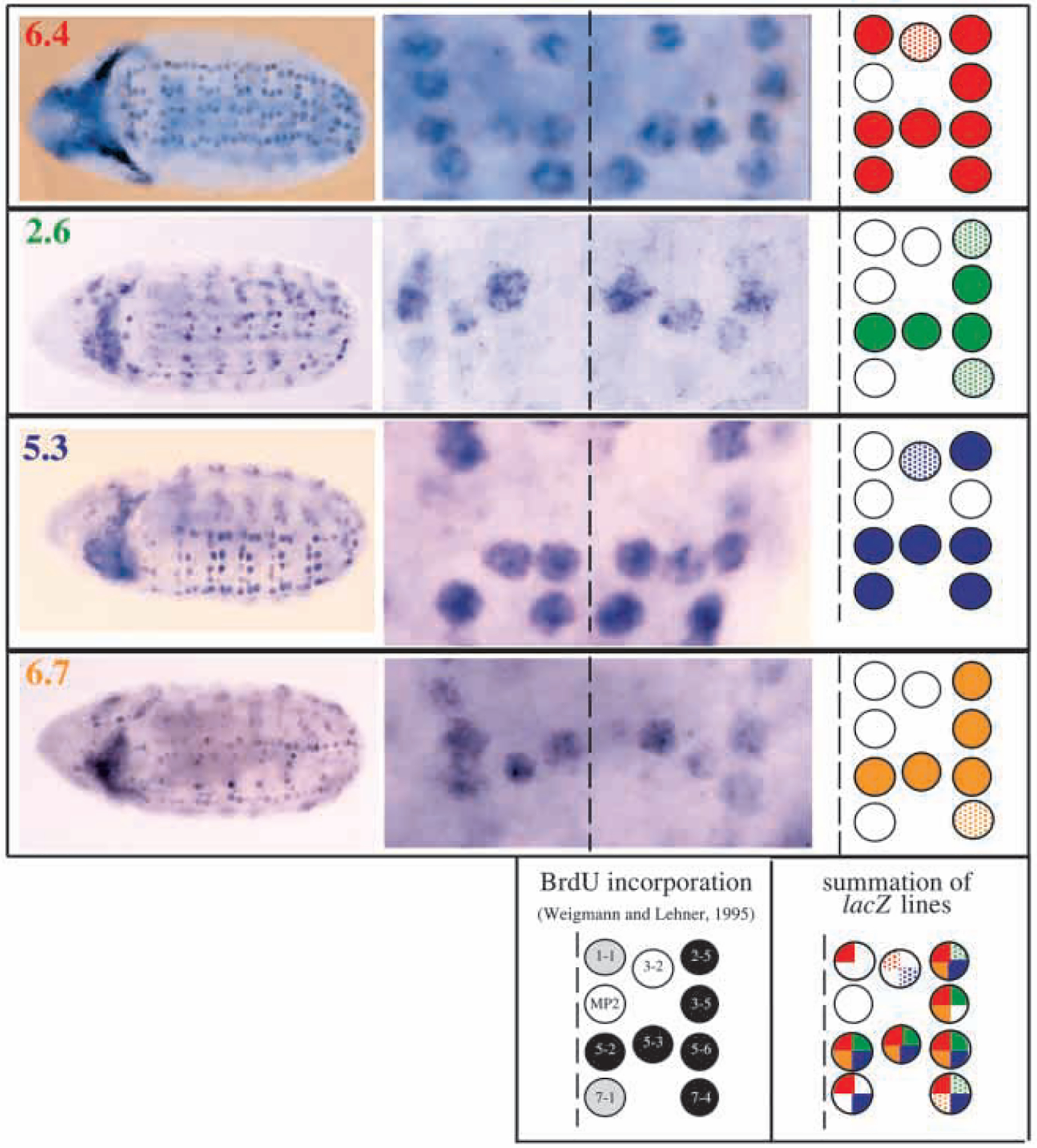
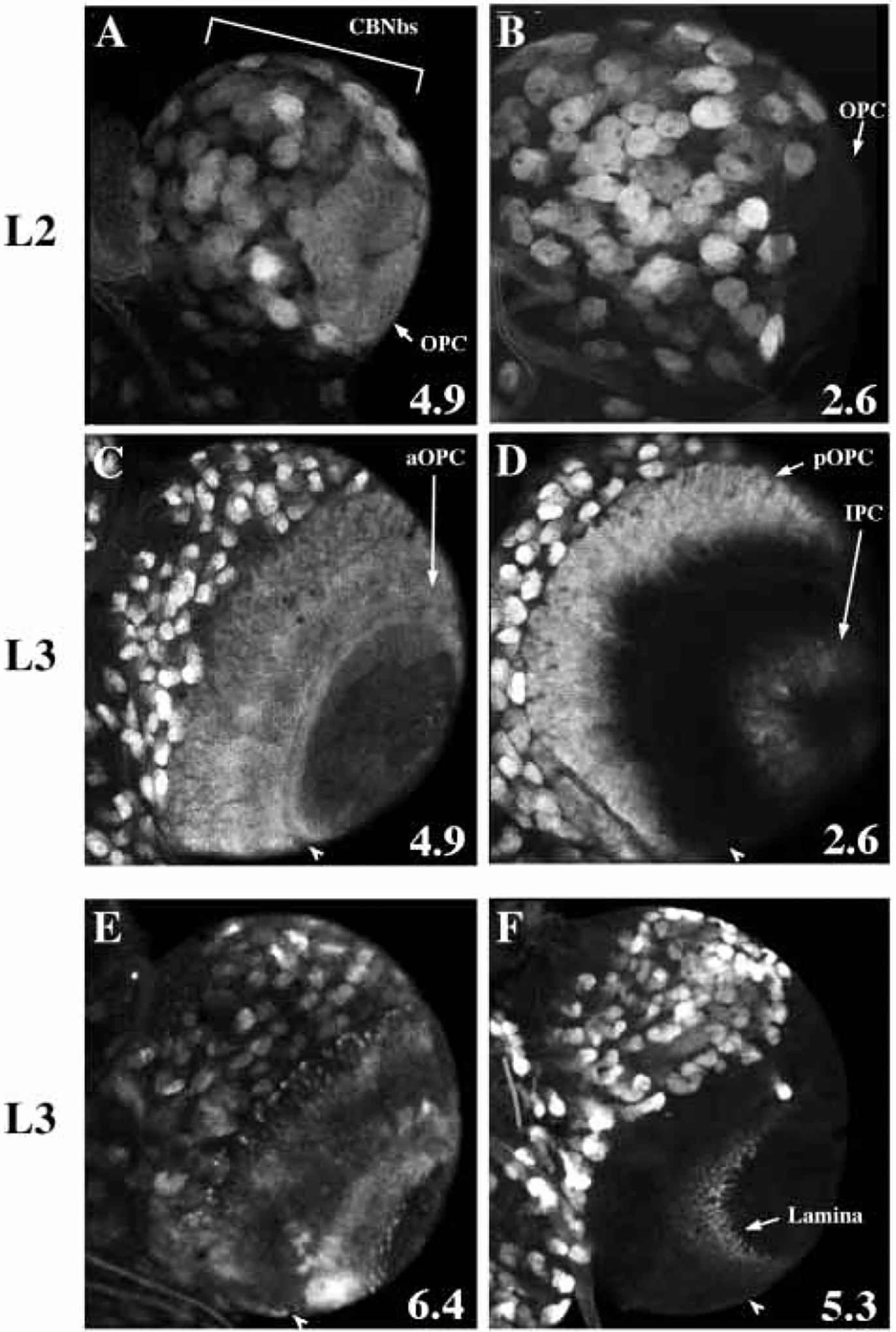
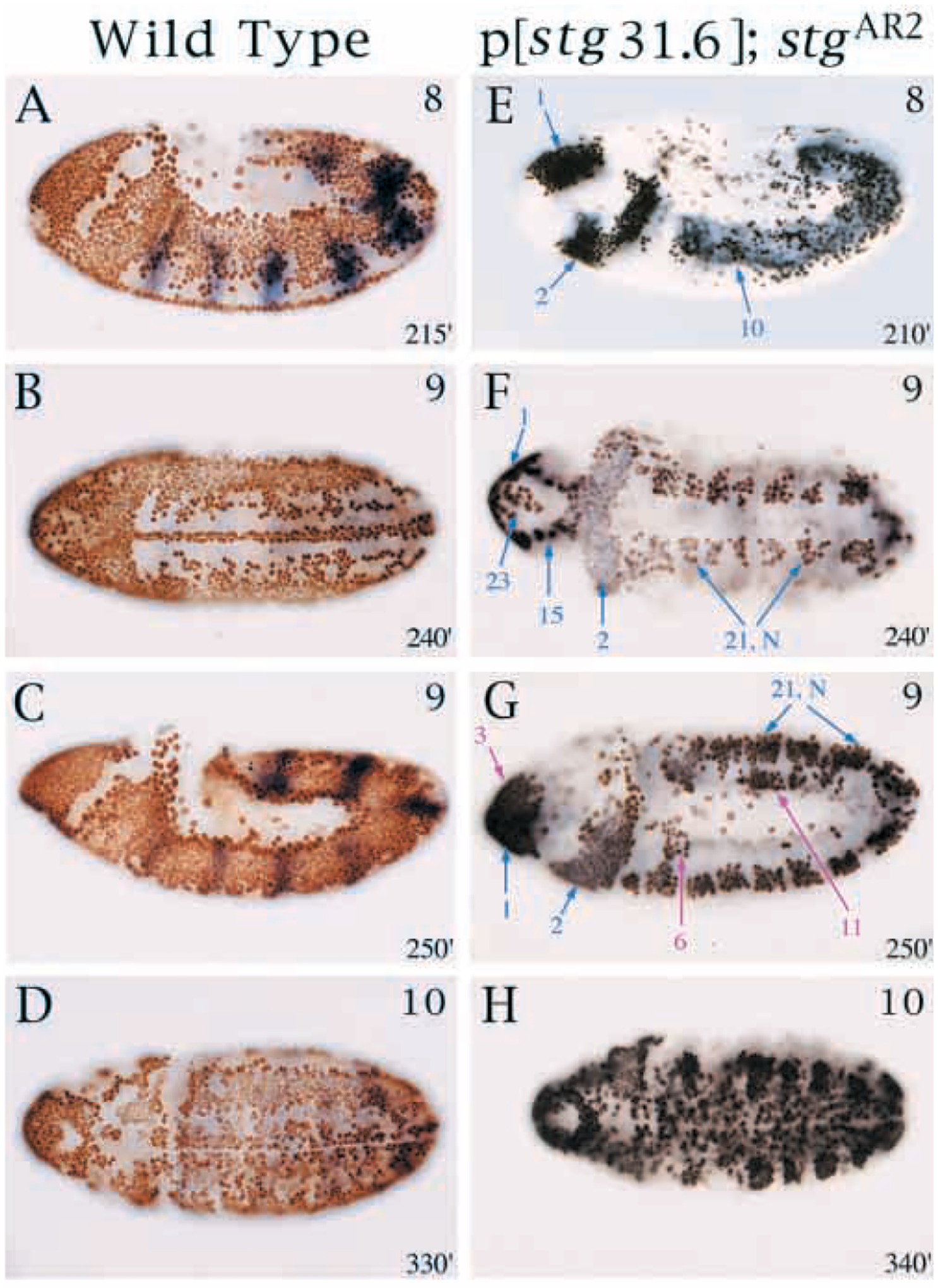
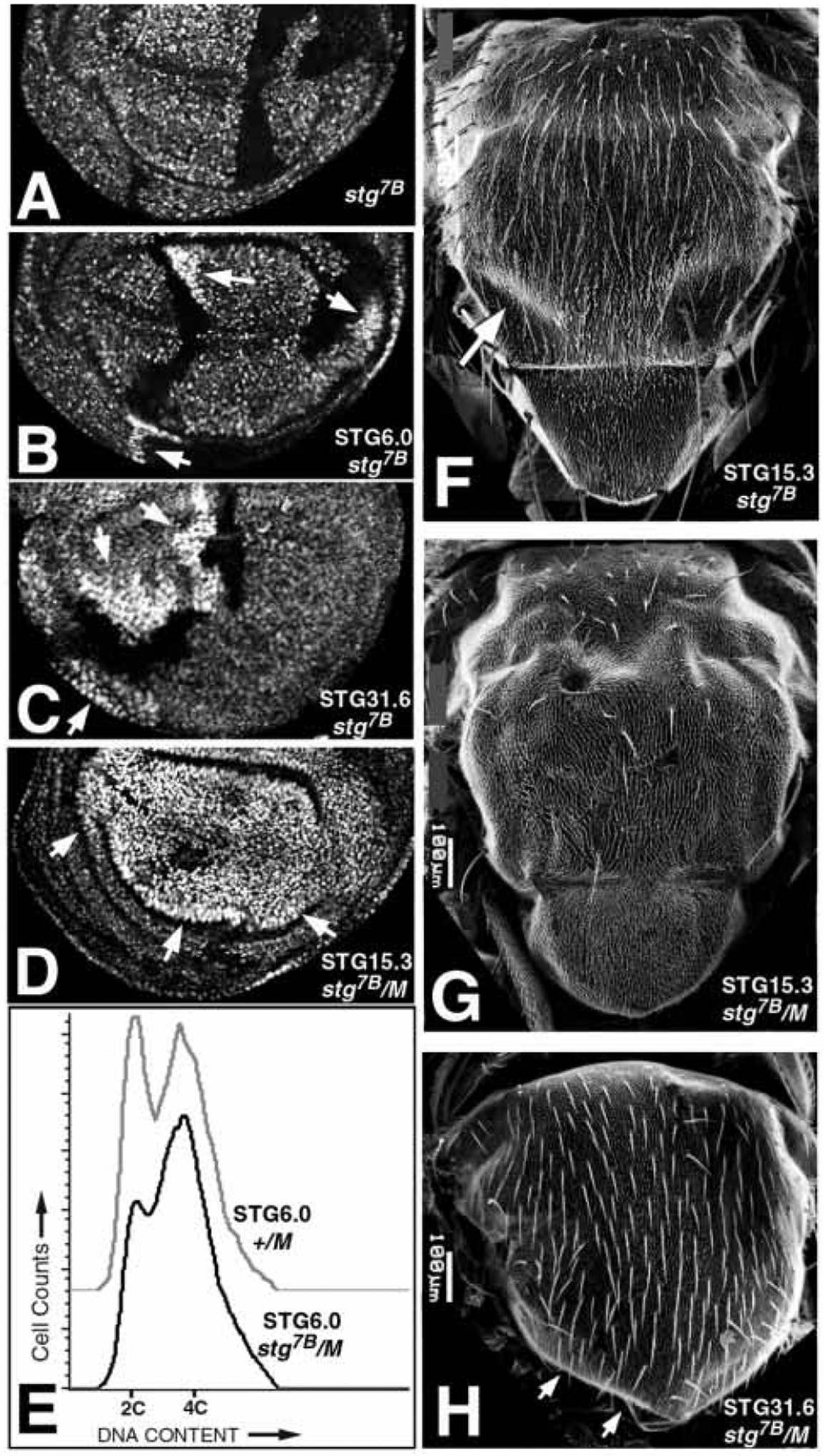
Mitosis in most Drosophila cells is triggered by brief bursts of transcription of string (stg), a Cdc25-type phosphatase that activates the mitotic kinase, Cdk1 (Cdc2). To understand how string transcription is regulated, we analyzed the expression of string-lacZ reporter genes covering approximately 40 kb of the string locus. We also tested protein coding fragments of the string locus of 6 kb to 31.6 kb for their ability to complement loss of string function in embryos and imaginal discs. A plethora of cis-acting elements spread over >30 kb control string transcription in different cells and tissue types. Regulatory elements specific to subsets of epidermal cells, mesoderm, trachea and nurse cells were identified, but the majority of the string locus appears to be devoted to controlling cell proliferation during neurogenesis. Consistent with this, compact promotor-proximal sequences are sufficient for string function during imaginal disc growth, but additional distal elements are required for the development of neural structures in the eye, wing, leg and notum. We suggest that, during evolution, cell-type-specific control elements were acquired by a simple growth-regulated promoter as a means of coordinating cell division with developmental processes, particularly neurogenesis.
Figures
References
-
- Arora K and Nüsslein-Volhard C (1992). Altered mitotic domains reveal fate map changes in Drosophila embryos mutant for zygotic dorsoventral patterning genes. Development 114, 1003–1024. - PubMed
-
- Bodmer R, Carretto R and Jan YN (1989). Neurogenesis of the peripheral nervous system in Drosophila embryos: DNA replication patterns and cell lineages. Neuron 3, 21–32. - PubMed
-
- Breeden L (1996). Start-specific transcription in yeast. Curr. Topics Microbiol. Immunol 208, 95–127. - PubMed
-
- Campos-Ortega JA and Hartenstein V (1985). The Embryonic Development of Drosophila melanogaster Berlin: Springer-Verlag.
-
- Crozatier M, Valle D, Dubois L, Ibnsouda S and Vincent A (1996). collier, a novel regulator of Drosophila head development, is expressed in a single mitotic domain. Curr. Biol 6, 707–718. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
