

Hypothalamic hypocretin (orexin): robust innervation of the spinal cord
- PMID: 10191330
- PMCID: PMC6782271
- DOI: 10.1523/JNEUROSCI.19-08-03171.1999
Hypothalamic hypocretin (orexin): robust innervation of the spinal cord
Abstract
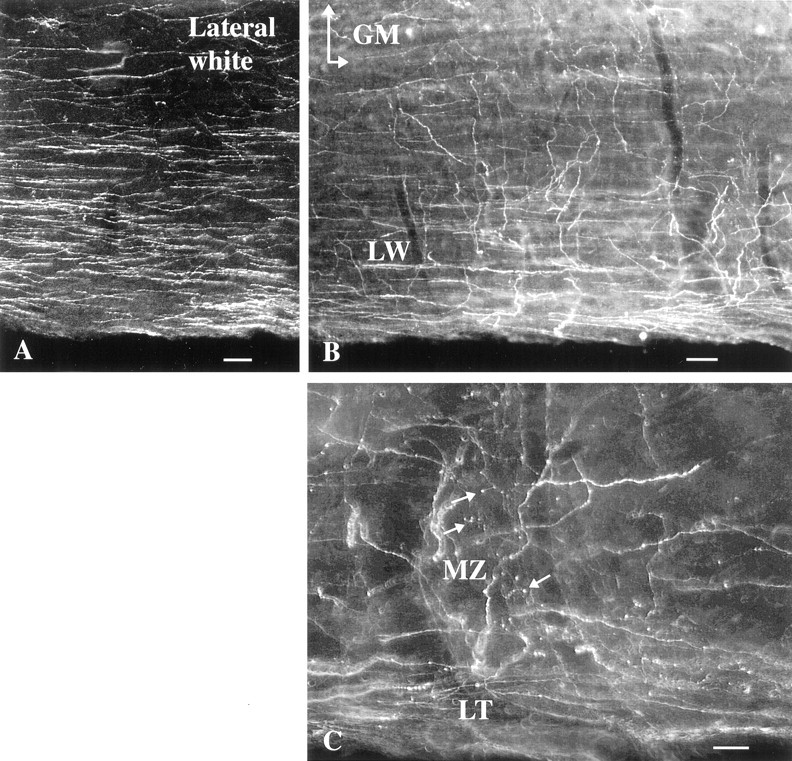
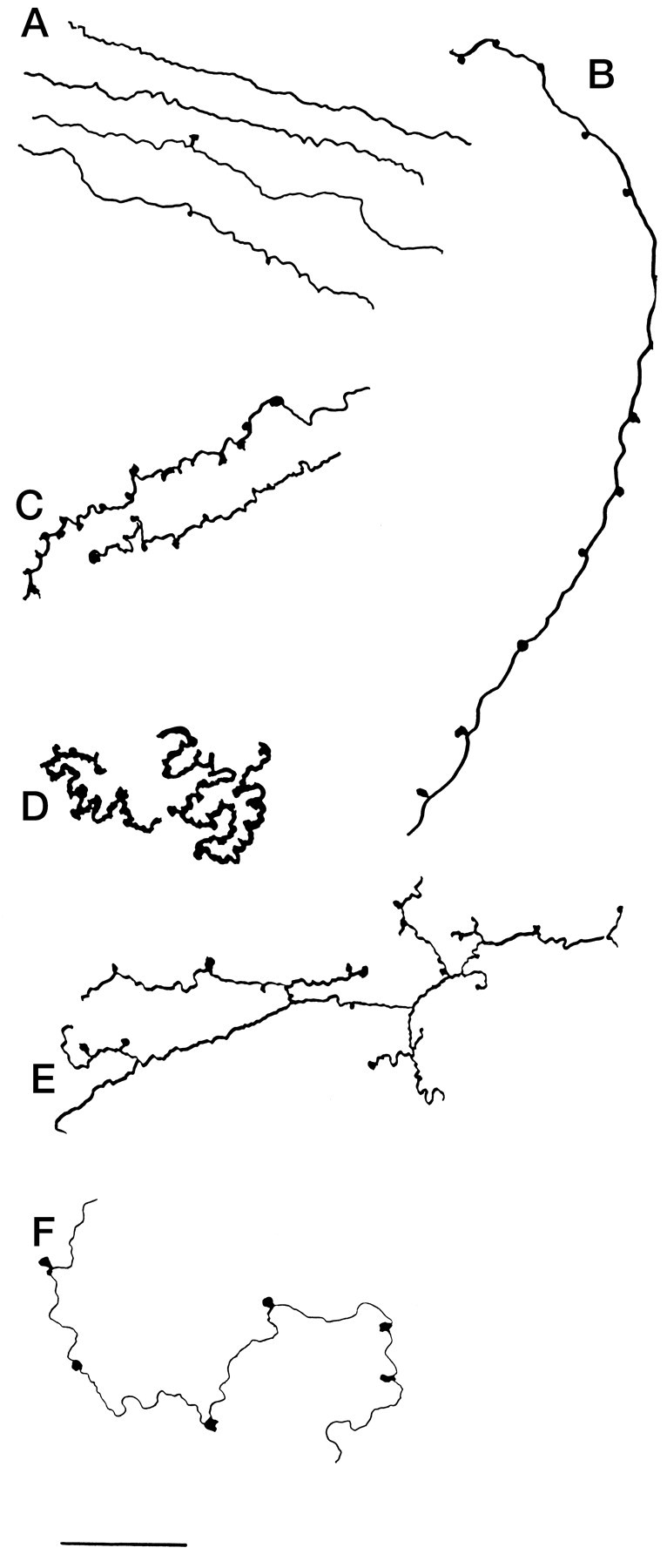
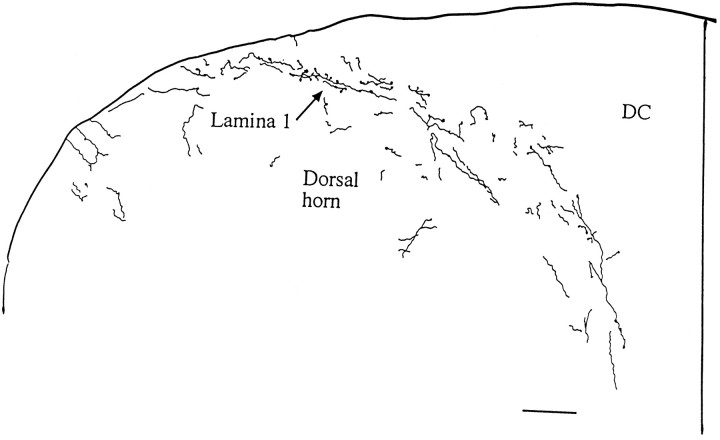
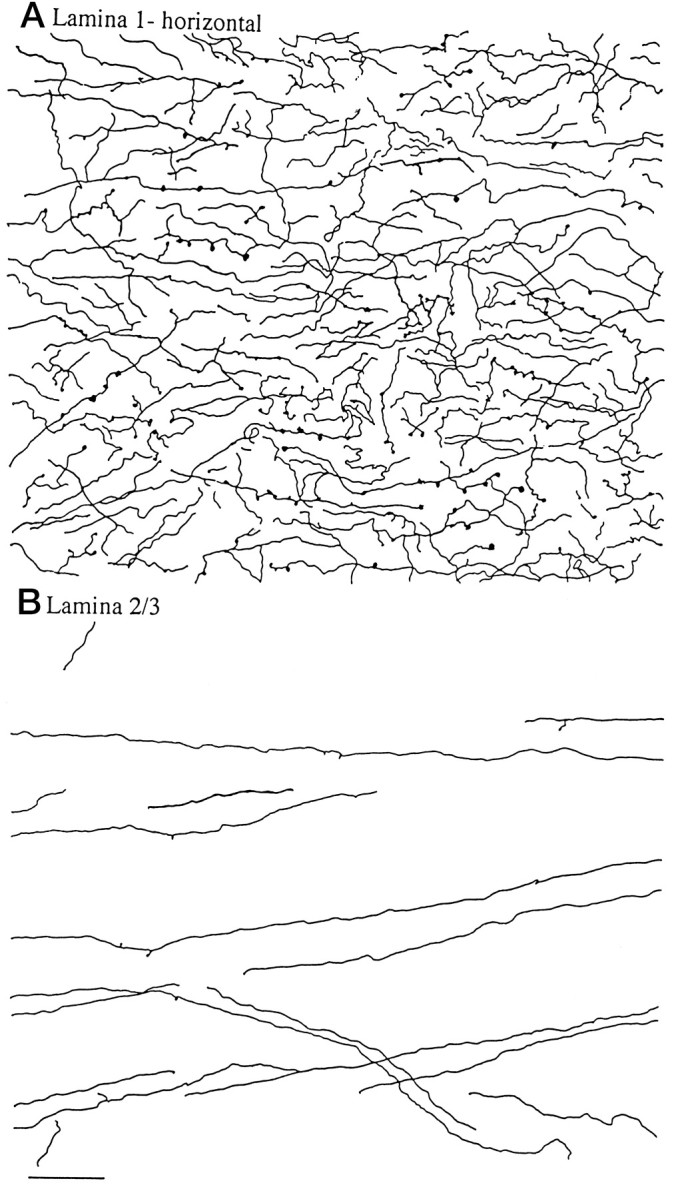
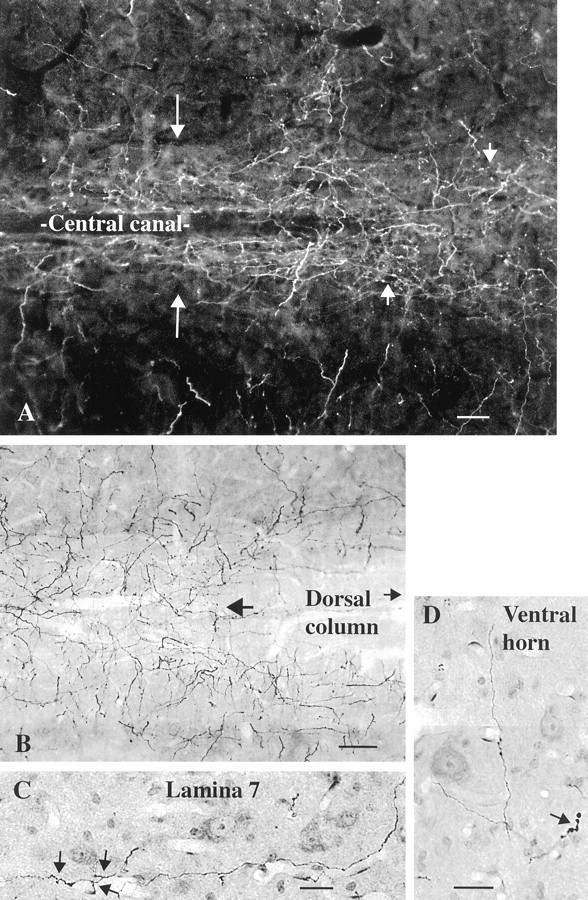
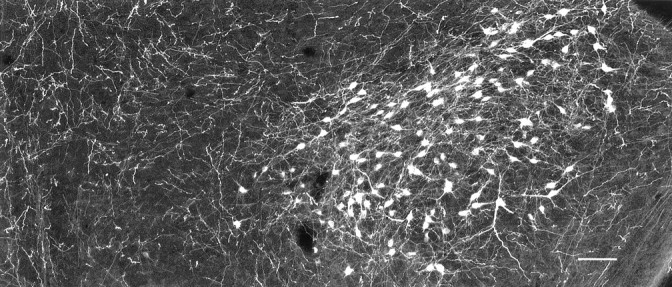
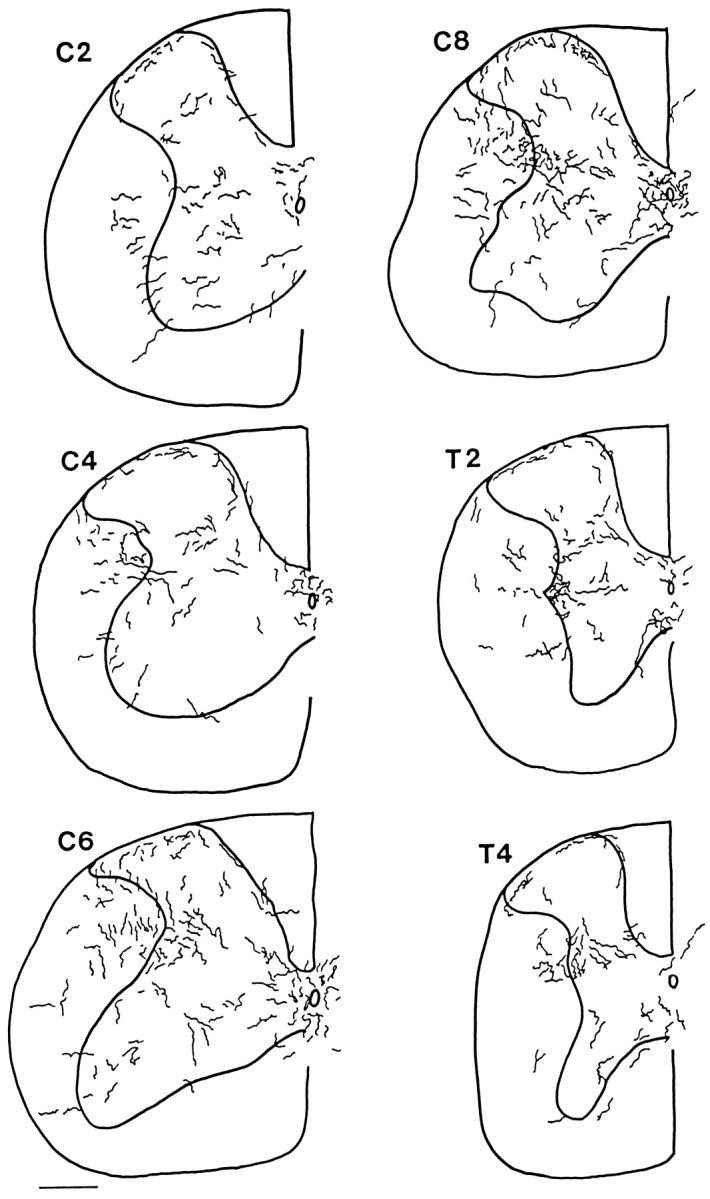
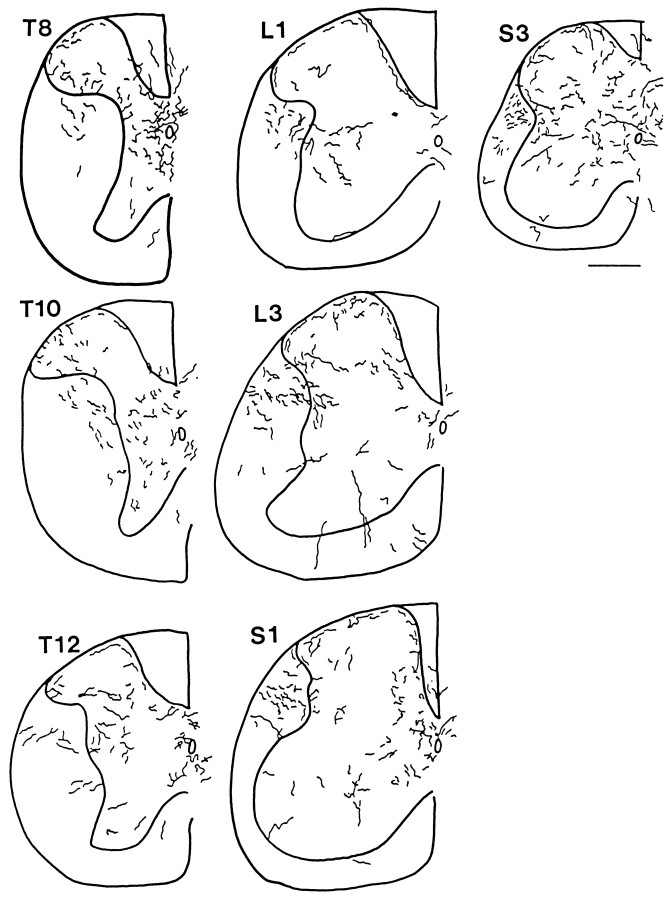
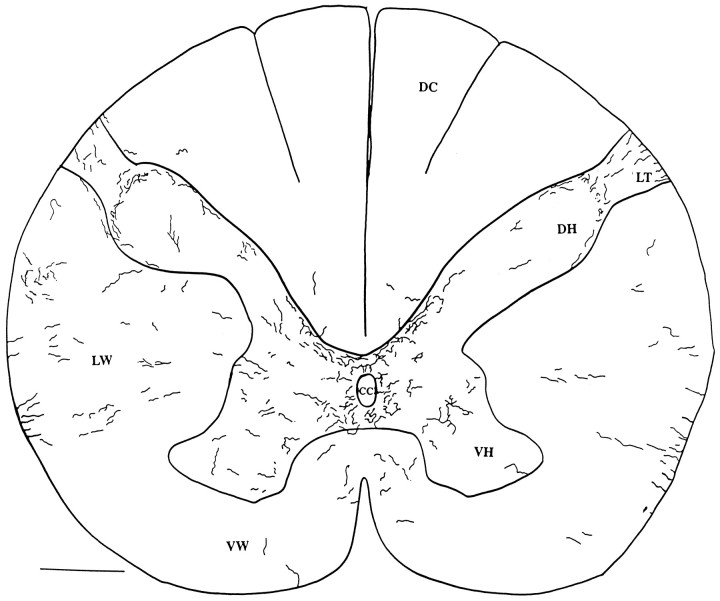
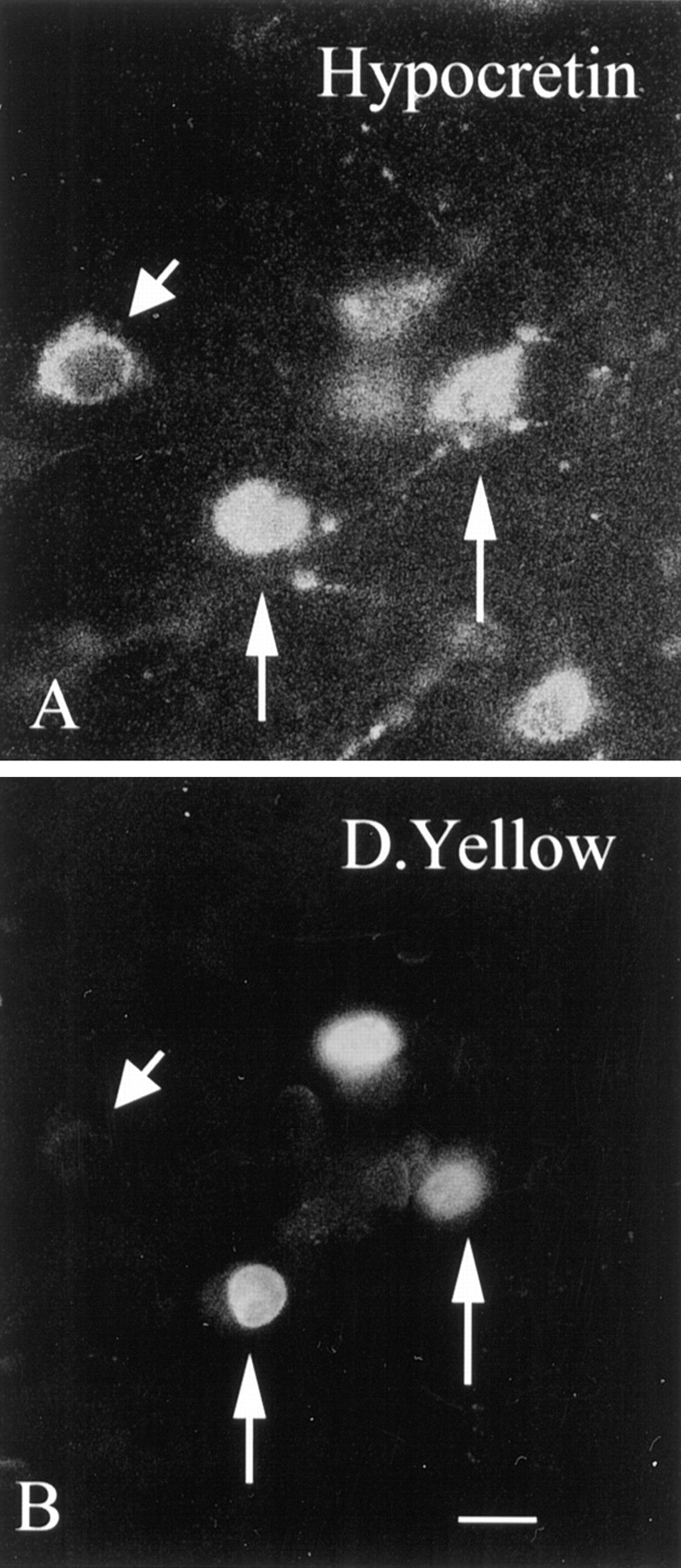
Hypocretin (orexin) is synthesized by neurons in the lateral hypothalamus and has been reported to increase food intake and regulate the neuroendocrine system. In the present paper, long descending axonal projections that contain hypocretin were found that innervate all levels of the spinal cord from cervical to sacral segments, as studied in mouse, rat, and human spinal cord and not previously described. High densities of axonal innervation are found in regions of the spinal cord related to modulation of sensation and pain, notably in the marginal zone (lamina 1). Innervation of the intermediolateral column and lamina 10 as well as strong innervation of the caudal region of the sacral cord suggest that hypocretin may participate in the regulation of both the sympathetic and parasympathetic parts of the autonomic nervous system. Double-labeling experiments in mice combining retrograde transport of diamidino yellow after spinal cord injections and immunocytochemistry support the concept that hypocretin-immunoreactive fibers in the cord originate from the neurons in the lateral hypothalamus. Digital-imaging physiological studies with fura-2 detected a rise in intracellular calcium in response to hypocretin in cultured rat spinal cord neurons, indicating that spinal cord neurons express hypocretin-responsive receptors. A greater number of cervical cord neurons responded to hypocretin than another hypothalamo-spinal neuropeptide, oxytocin. These data suggest that in addition to possible roles in feeding and endocrine regulation, the descending hypocretin fiber system may play a role in modulation of sensory input, particularly in regions of the cord related to pain perception and autonomic tone.
Figures


References
-
- Armstrong WE, Warach S, Hatton GI, NcNeill TH. Subnuclei in the rat paraventricular nucleus: a cytoarcitectural, HRP, and immunocytochemical analysis. Neuroscience. 1980;206:317–345. - PubMed
-
- Bittencourt JC, Elias CF. Melanin-concentrating hormone and neuropeptide EI projections from the lateral hypothalamic area and zona incerta to the medial septal nucleus and spinal cord: a study using multiple neuronal tracers. Brain Res. 1998;805:1–19. - PubMed
-
- Bittencourt JC, Presse F, Arias C, Vaughan J, Nahon JL, Vale W, Sawchenko PE. The melanin-concentrating hormone system of the rat brain: an immuno- and hybridization histochemical characterization. J Comp Neurol. 1992;319:218–245. - PubMed
-
- Brown AG. Organization in the spinal cord. The anatomy and physiology of identified neurons, pp 1–238. Springer; New York: 1981.
-
- Cedarbaum JM, Aghajanian GK. Afferent projections to the rat locus coeruleus as determined by a retrograde tracing technique. J Comp Neurol. 1978;178:1–16. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources