

Ordering of ceramide formation, caspase activation, and mitochondrial changes during CD95- and DNA damage-induced apoptosis
- PMID: 10194469
- PMCID: PMC408258
- DOI: 10.1172/JCI5457
Ordering of ceramide formation, caspase activation, and mitochondrial changes during CD95- and DNA damage-induced apoptosis
Erratum in
- J Clin Invest 1999 May;103(9):1363
Abstract
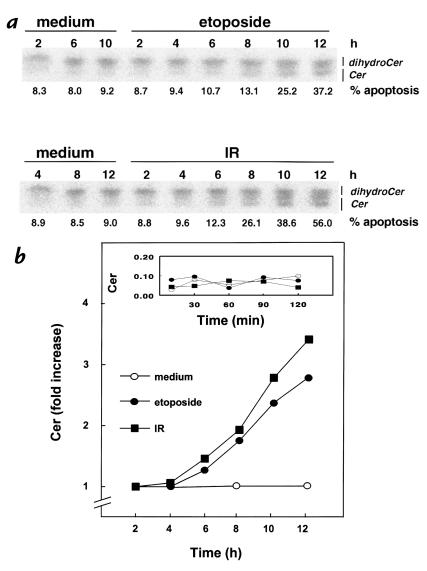
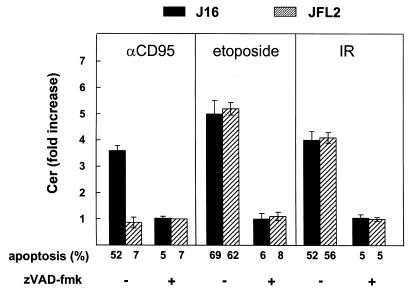
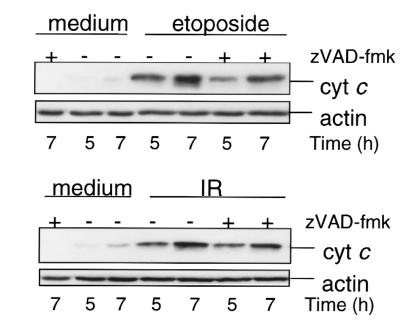
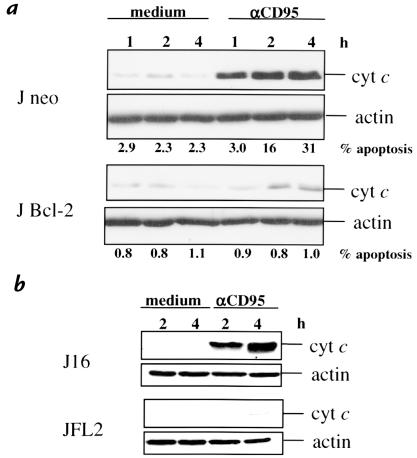
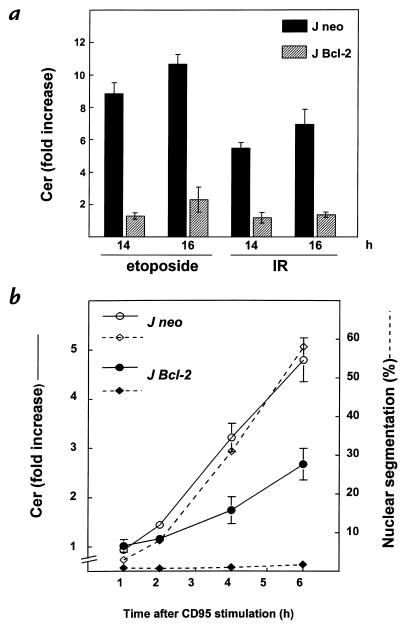
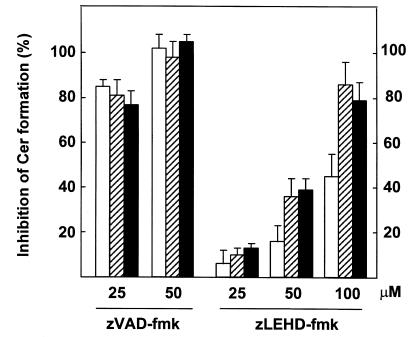
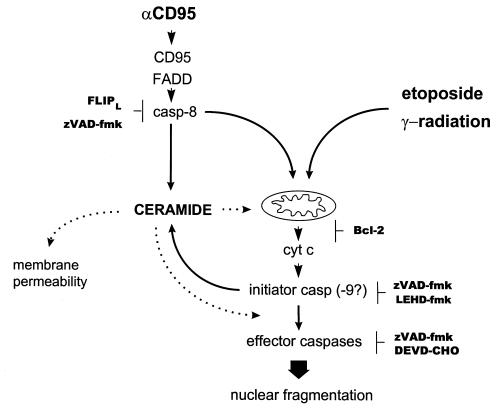
To evaluate the role of ceramide (Cer) in apoptosis signaling, we examined Cer formation induced by CD95, etoposide, or gamma-radiation (IR) in relation to caspase activation and mitochondrial changes in Jurkat T cells. The Cer response to all three stimuli was mapped in between caspases sensitive to benzoyloxycarbonyl-VAD-fluoromethylketone (zVAD-fmk) and acetyl-DEVD-aldehyde (DEVD-CHO). Cer production was independent of nuclear fragmentation but associated with the occurrence of other aspects of the apoptotic morphology. Caspase-8 inhibition abrogated Cer formation and apoptosis induced by CD95 but did not affect the response to etoposide or IR, placing CD95-induced Cer formation downstream from caspase-8 and excluding a role for caspase-8 in the DNA damage pathways. CD95 signaling to the mitochondria required caspase-8, whereas cytochrome c release in response to DNA damage was caspase-independent. These results indicate that the caspases required for the Cer response to etoposide and IR reside at or downstream from the mitochondria. Bcl-2 overexpression abrogated the Cer response to etoposide and IR and reduced CD95-induced Cer accumulation. We conclude that the Cer response to DNA damage fully depends on mitochondrion-dependent caspases, whereas the response to CD95 partially relies on these caspases. Our data imply that Cer is not instrumental in the activation of inducer caspases or signaling to the mitochondria. Rather, Cer formation is associated with the execution phase of apoptosis.
Figures
References
-
- Hannun YA. Functions of ceramide in coordinating cellular responses to stress. Science. 1996;274:1855–1859. - PubMed
-
- Wiegmann K, Schutze S, Machleidt T, Witte D, Kronke M. Functional dichotomy of neutral and acidic sphingomyelinases in tumor necrosis factor signaling. Cell. 1994;78:1005–1015. - PubMed
-
- Santana P, et al. Acid sphingomyelinase-deficient human lymphoblasts and mice are defective in radiation-induced apoptosis. Cell. 1996;86:189–199. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
