

Proteolysis of monocyte CD14 by human leukocyte elastase inhibits lipopolysaccharide-mediated cell activation
- PMID: 10194477
- PMCID: PMC408261
- DOI: 10.1172/JCI5779
Proteolysis of monocyte CD14 by human leukocyte elastase inhibits lipopolysaccharide-mediated cell activation
Abstract
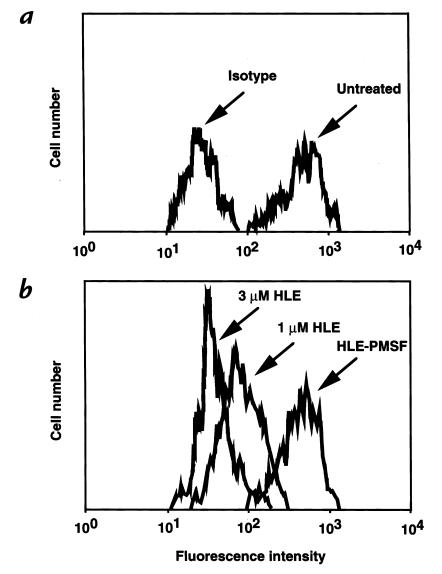
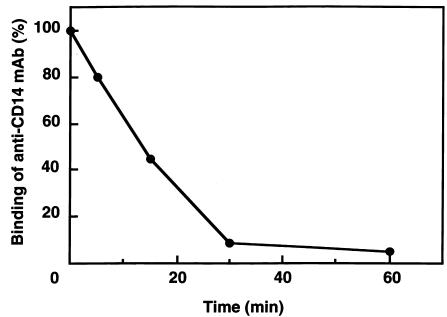
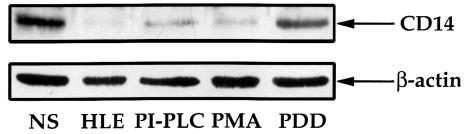
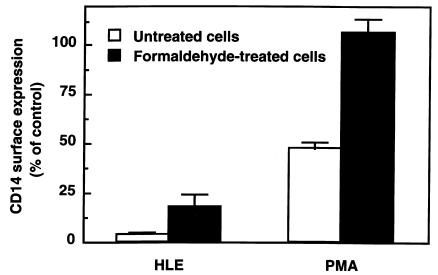
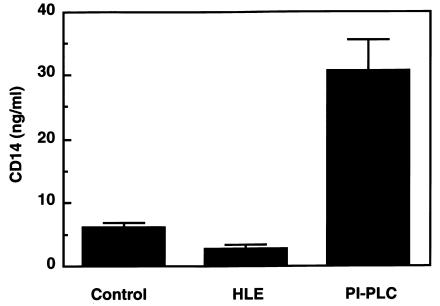
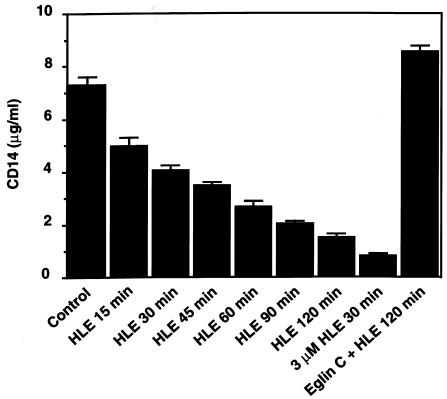
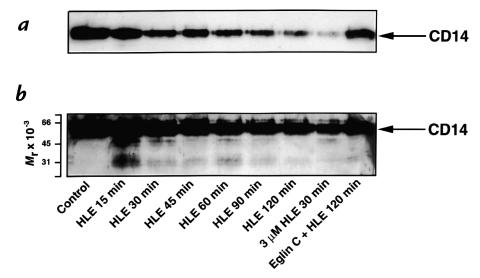
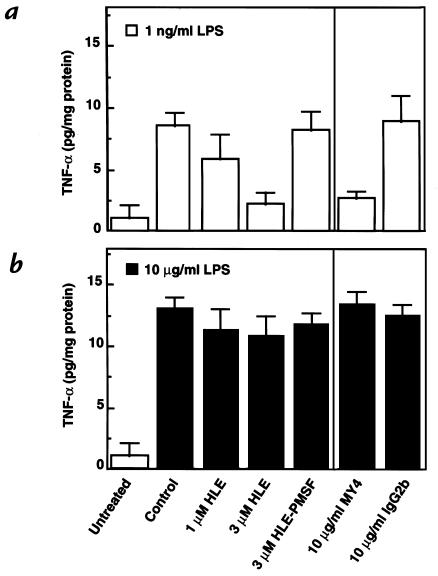
Human leukocyte elastase (HLE), a polymorphonuclear neutrophil (PMN) serine proteinase, is proteolytically active on some membrane receptors at the surface of immune cells. The present study focused on the effect of HLE on the expression of CD14, the main bacterial lipopolysaccharide (LPS) receptor at the surface of monocytes. HLE exhibited a time- and concentration-dependent downregulatory effect on CD14 surface expression. A 30-minute incubation of 3 microM HLE was required to display 95% disappearance of the receptor. This downregulation resulted from a direct proteolytic process, not from a shedding consecutive to monocyte activation as observed upon challenge with phorbol myristate acetate (PMA). To confirm that CD14 is a substrate for HLE, this enzyme was incubated with recombinant human CD14 (Mr approximately 57,000), and proteolysis was further analyzed by immunoblot analysis. Cleavage of the CD14 molecule was directly evidenced by the generation of short-lived fragments (Mr approximately 47,000 and 30,000). As a consequence of the CD14 proteolysis, a decrease in the responsiveness of monocytes to LPS was observed, as assessed by measuring tumor necrosis factor-alpha (TNF-alpha) formation. This inhibition was only observed with 1 ng/ml of LPS, i.e., when only the CD14-dependent pathway was involved. At a higher LPS concentration, such as 10 microgram/ml, when CD14-independent pathways were operative, this inhibition was overcome. The direct proteolysis by HLE of the membrane CD14 expressed on monocytes illustrates a potential anti-inflammatory effect of HLE through inhibition of LPS-mediated cell activation.
Figures
References
-
- Smith JA. Neutrophils, host defense, and inflammation: a double-edged sword. J Leukoc Biol. 1994;56:672–686. - PubMed
-
- Woessner, J.F., Jr. 1992. Role of cellular proteinases and their protein inhibitors in inflammation. In Biochemistry of inflammation. J.T. Whicher and S.W. Evans, editors. Kluwer Academic Publishers. Lancaster, PA. 57–89.
-
- Döring G. The role of neutrophil elastase in chronic inflammation. Am J Respir Crit Care Med. 1994;150:S114–S117. - PubMed
-
- Travis J, Pike R, Imamura T, Potempa J. The role of proteolytic enzymes in the development of pulmonary emphysema and periodontal disease. Am J Respir Crit Care Med. 1994;150:S143–S146. - PubMed
-
- Bieth, J.G. 1986. Elastases: catalytic and biological properties. In Regulation of matrix accumulation. R. Mecham, editor. Academic Press. New York, NY. 217–230.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
