

doi: 10.1101/gad.13.7.792.
Toll receptor-mediated Drosophila immune response requires Dif, an NF-kappaB factor
Affiliations
- PMID: 10197979
- PMCID: PMC316597
- DOI: 10.1101/gad.13.7.792
Item in Clipboard
Toll receptor-mediated Drosophila immune response requires Dif, an NF-kappaB factor
Genes Dev.
.
Abstract
The induction of immunity genes in Drosophila has been proposed to be dependent on Dorsal, Dif, and Relish, the NF-kappaB-related factors. Here we provide genetic evidence that Dif is required for the induction of only a subset of antimicrobial peptide genes. The results show that the presence of Dif without Dorsal is sufficient to mediate the induction of drosomycin and defensin. We also demonstrate that Dif is a downstream component of the Toll signaling pathway in activating the drosomycin expression. These results reveal that individual members of the NF-kappaB family in Drosophila have distinct roles in immunity and development.
Figures
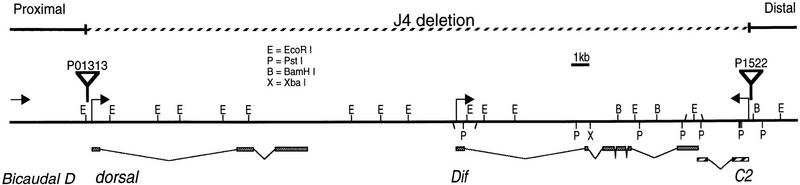
Restriction map and intron–exon structure of the Dif–dorsal locus, which includes dorsal, Dif and C2. Bicaudal D is located ∼5 kb upstream of dorsal (Wharton and Struhl 1989). Arrows indicate the start sites and directions of transcription, except the arrow on Bicaudal D shows only the direction of transcription. The exons are represented by rectangular boxes; the introns are represented by the angled lines. All the known EcoRI restriction sites are shown; PstI, BamHI, and XbaI sites are shown only on the Dif and C2 genes. The EcoRI restriction map of the dorsal gene is duplicated from Steward (1987). P-element insertions are indicated by flags. The deletion in the J4 chromosome (broken line), uncovers the three genes but does not affect Bicaudal D.
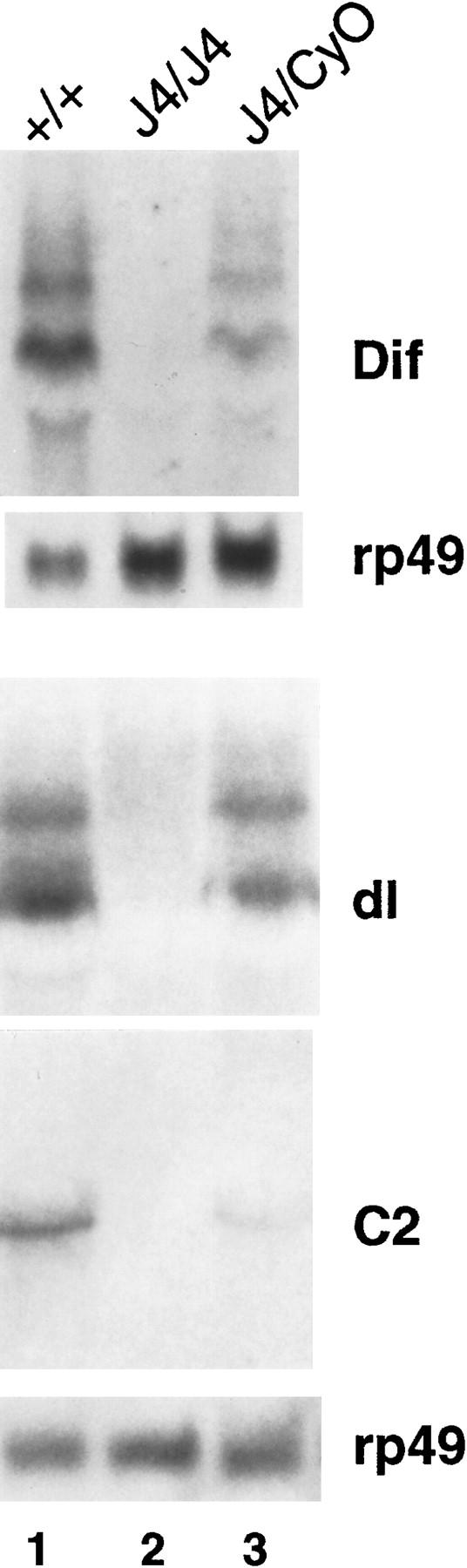
Northern analysis of the expression of the Dif–dorsal locus in the J4 deletion. Total RNAs were isolated from Canton-S wild-type flies (+/+ , lane 1), J4 homozygous flies (J4/J4, lane 2), or heterozygous J4 flies (J4/CyO, lane 3). RNAs were analyzed on denaturing gels, transferred to membranes, and hybridized with radiolabeled probes as indicated (right). (dl) dorsal. The ribosomal protein gene rp49 was used as a loading control.
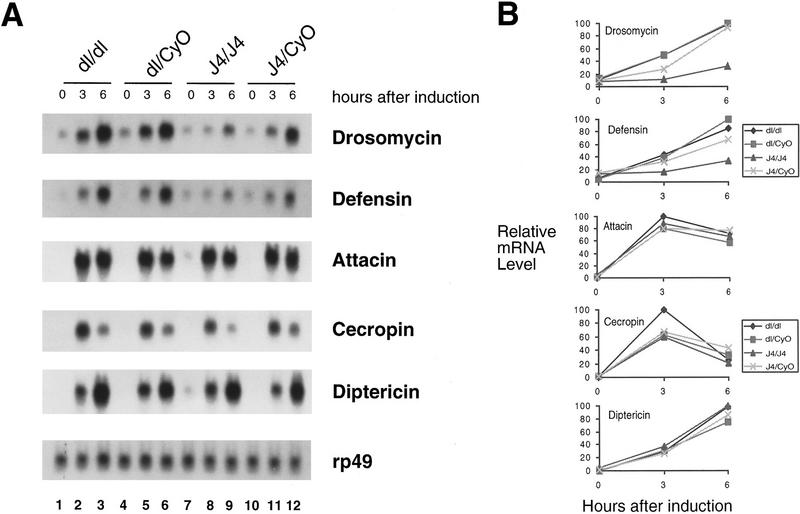
Induction of antimicrobial peptide genes in the Dif/dorsal deletion mutant. (A) Total RNAs were isolated from fly strains with the genotype indicated at top. The dorsal (dl) mutant used in this and other experiments was the parental P01313 line. Prior to RNA isolation, the flies were induced by injection of E. coli bacteria and allowed to recover for 3 or 6 hr. The 0-hour represents no injection. The isolated RNAs were analyzed by Northern blot and hybridized with radiolabeled probes indicated at right. Each panel utilized the same set of RNA isolated in parallel, but the times of exposure varied from 12 to 72 hr. This is a representative result of three independent experiments. (B) Quantitative analysis of gene induction. The hybridization signal of the experiments shown in A was quantitated using PhosphorImager (Molecular Dynamics). The relative mRNA levels, normalized with the rp49 signal, were plotted as shown. The value of 100 was assigned to the highest signals of the individual blots; the other signals were calculated as a fraction of this value.
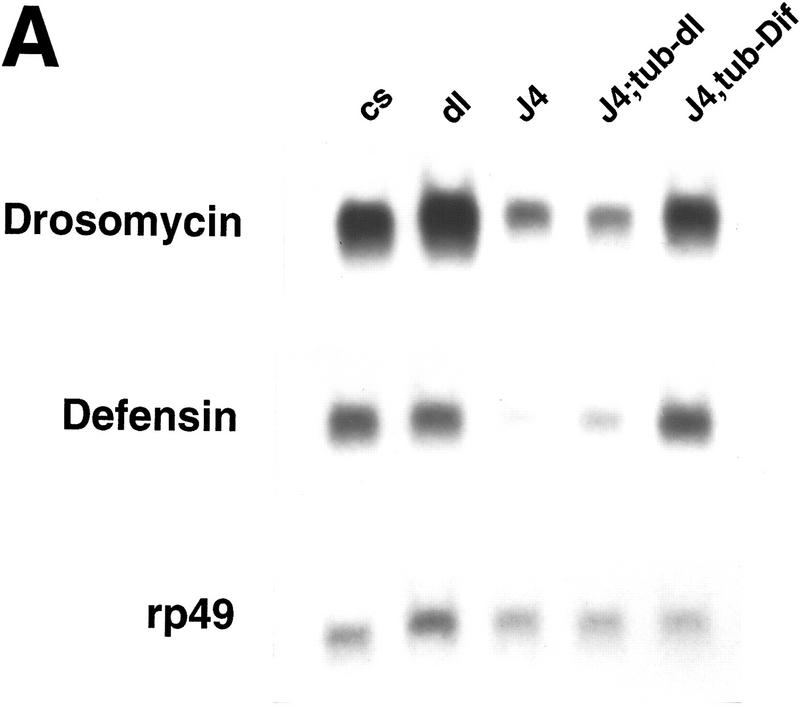
Ubiquitous expression of Dif rescues the defect of immune response. (A) Transgenic flies containing the tubulinα1–dorsal or -Dif constructs were crossed or recombined, respectively, with the J4 chromosome. Flies that had the genotypes indicated were induced by bacterial injection and after 6 hr the RNA were isolated for Northern analysis. The autoradiographs of the blots hybridized with the indicated probes are shown. (B) The blots were quantitated using PhosphorImager, and the drosomycin and defensin signals were normalized with that of rp49. The relative levels of mRNA expression were plotted, with the Canton S (cs) wild-type fly expression levels assigned as 1. The graphs show the average result of two independent experiments. (C) Control experiments were carried out using the transgenic lines to examine the expression of dorsal in adult flies. The tubulin–dorsal transgene drives the expression of dorsal mRNA to a level higher than that in the parental y w flies. (D) The severe cuticle phenotype exhibited in embryos derived from dorsalI5 mutant mothers (left) could be rescued by the tubulin-dorsal transgene (right), demonstrating that the transgene can produce functional Dorsal proteins.
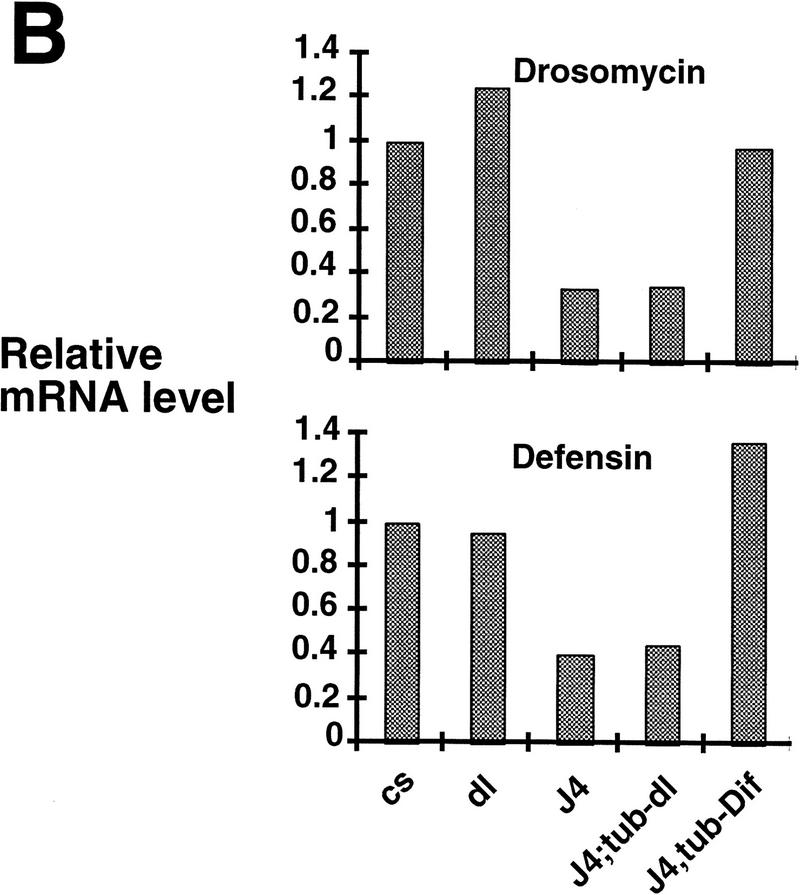
Ubiquitous expression of Dif rescues the defect of immune response. (A) Transgenic flies containing the tubulinα1–dorsal or -Dif constructs were crossed or recombined, respectively, with the J4 chromosome. Flies that had the genotypes indicated were induced by bacterial injection and after 6 hr the RNA were isolated for Northern analysis. The autoradiographs of the blots hybridized with the indicated probes are shown. (B) The blots were quantitated using PhosphorImager, and the drosomycin and defensin signals were normalized with that of rp49. The relative levels of mRNA expression were plotted, with the Canton S (cs) wild-type fly expression levels assigned as 1. The graphs show the average result of two independent experiments. (C) Control experiments were carried out using the transgenic lines to examine the expression of dorsal in adult flies. The tubulin–dorsal transgene drives the expression of dorsal mRNA to a level higher than that in the parental y w flies. (D) The severe cuticle phenotype exhibited in embryos derived from dorsalI5 mutant mothers (left) could be rescued by the tubulin-dorsal transgene (right), demonstrating that the transgene can produce functional Dorsal proteins.
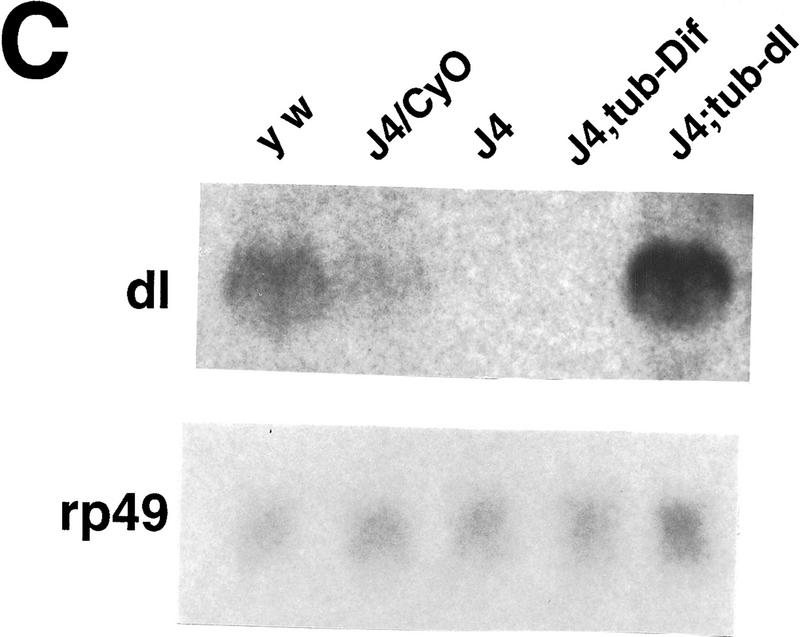
Ubiquitous expression of Dif rescues the defect of immune response. (A) Transgenic flies containing the tubulinα1–dorsal or -Dif constructs were crossed or recombined, respectively, with the J4 chromosome. Flies that had the genotypes indicated were induced by bacterial injection and after 6 hr the RNA were isolated for Northern analysis. The autoradiographs of the blots hybridized with the indicated probes are shown. (B) The blots were quantitated using PhosphorImager, and the drosomycin and defensin signals were normalized with that of rp49. The relative levels of mRNA expression were plotted, with the Canton S (cs) wild-type fly expression levels assigned as 1. The graphs show the average result of two independent experiments. (C) Control experiments were carried out using the transgenic lines to examine the expression of dorsal in adult flies. The tubulin–dorsal transgene drives the expression of dorsal mRNA to a level higher than that in the parental y w flies. (D) The severe cuticle phenotype exhibited in embryos derived from dorsalI5 mutant mothers (left) could be rescued by the tubulin-dorsal transgene (right), demonstrating that the transgene can produce functional Dorsal proteins.
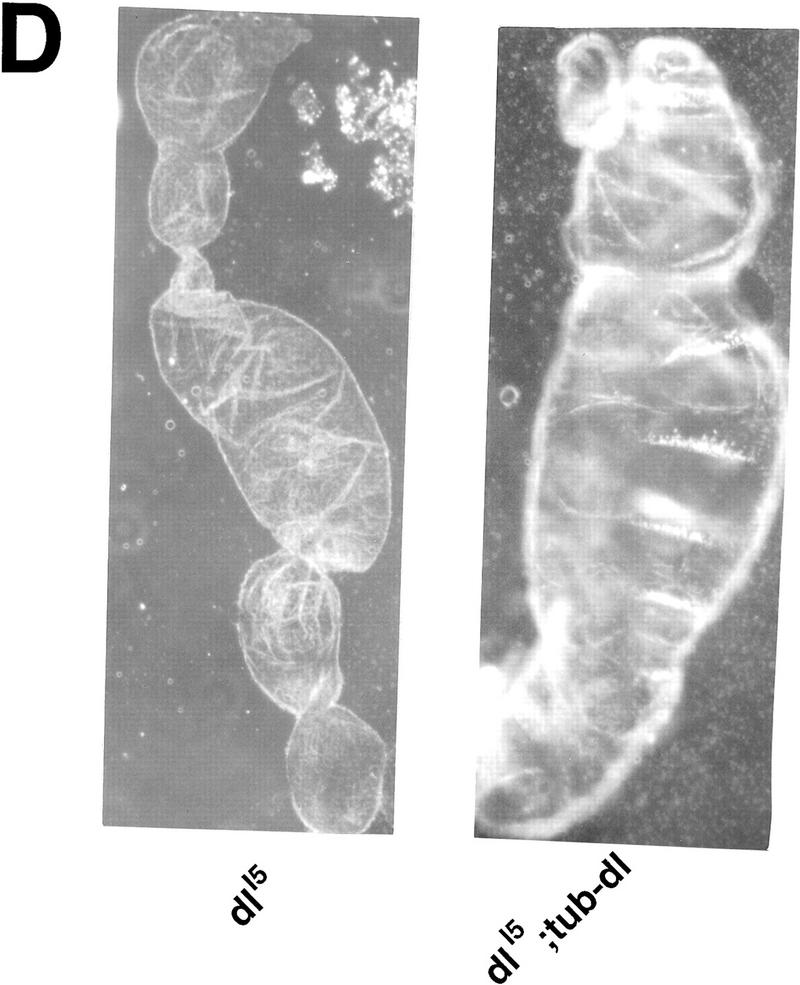
Ubiquitous expression of Dif rescues the defect of immune response. (A) Transgenic flies containing the tubulinα1–dorsal or -Dif constructs were crossed or recombined, respectively, with the J4 chromosome. Flies that had the genotypes indicated were induced by bacterial injection and after 6 hr the RNA were isolated for Northern analysis. The autoradiographs of the blots hybridized with the indicated probes are shown. (B) The blots were quantitated using PhosphorImager, and the drosomycin and defensin signals were normalized with that of rp49. The relative levels of mRNA expression were plotted, with the Canton S (cs) wild-type fly expression levels assigned as 1. The graphs show the average result of two independent experiments. (C) Control experiments were carried out using the transgenic lines to examine the expression of dorsal in adult flies. The tubulin–dorsal transgene drives the expression of dorsal mRNA to a level higher than that in the parental y w flies. (D) The severe cuticle phenotype exhibited in embryos derived from dorsalI5 mutant mothers (left) could be rescued by the tubulin-dorsal transgene (right), demonstrating that the transgene can produce functional Dorsal proteins.
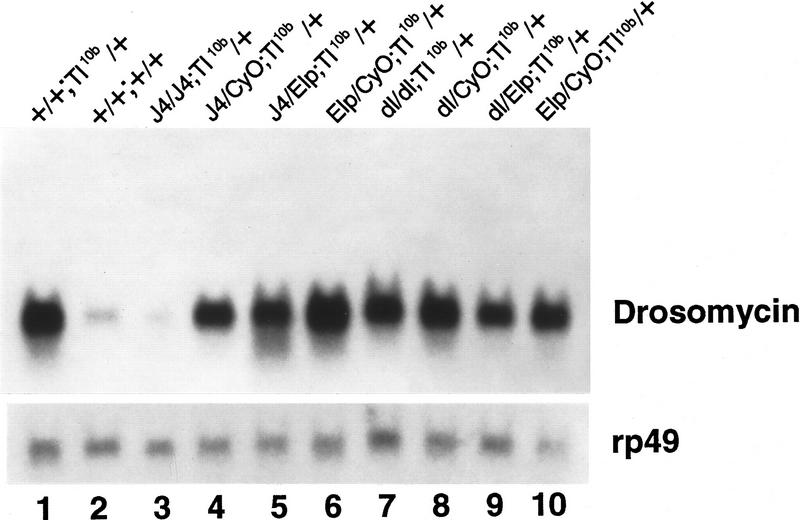
Dif is a downstream component of the Toll signaling pathway. Toll10b gain-of-function mutant male flies were mated with yw; BcElp/CyO; Ki/TM6y+ double balancer flies. Similarly, the J4/CyO or dl01313/CyO flies were mated with double balancer flies. F1 flies with the correct markers were mated together to generate the F2 flies (genotype shown above lanes 3–10). The flies were used directly without bacterial injection for RNA isolation and Northern analysis. An autoradiograph after hybridization with the drosomycin probe is shown.
References
-
- Attar RM, Caamano J, Carrasco D, Iotsova V, Ishikawa H, Ryseck RP, Weih F, Bravo R. Genetic approaches to study Rel/NF-kappaB/I kappaB function in mice. Semin Cancer Biol. 1997;8:93–101. - PubMed
-
- Baeuerle PA, Baltimore D. NF-κB: Ten years after. Cell. 1996;87:13–20. - PubMed
-
- Basler K, Struhl G. Compartment boundaries and the control of Drosophila limb pattern by hedgehog protein. Nature. 1994;368:208–214. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases