

Distinct domains within Vps35p mediate the retrieval of two different cargo proteins from the yeast prevacuolar/endosomal compartment
- PMID: 10198044
- PMCID: PMC25208
- DOI: 10.1091/mbc.10.4.875
Distinct domains within Vps35p mediate the retrieval of two different cargo proteins from the yeast prevacuolar/endosomal compartment
Abstract
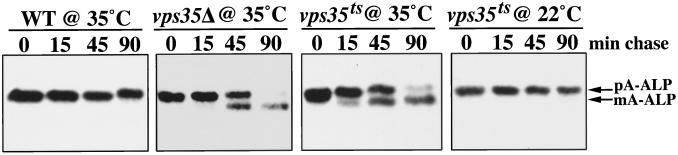
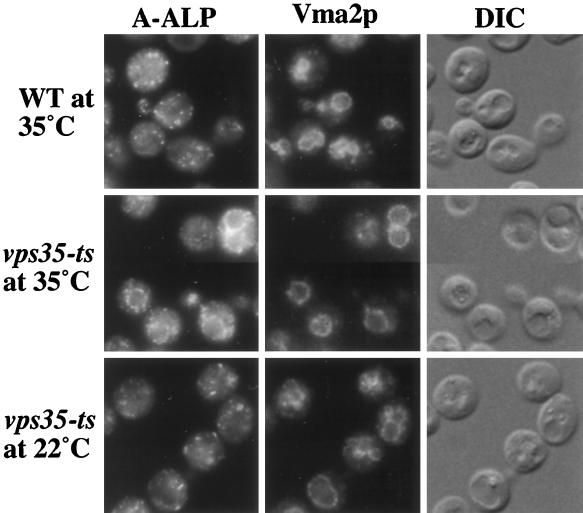
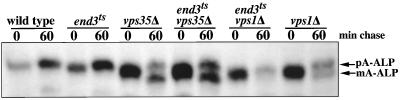
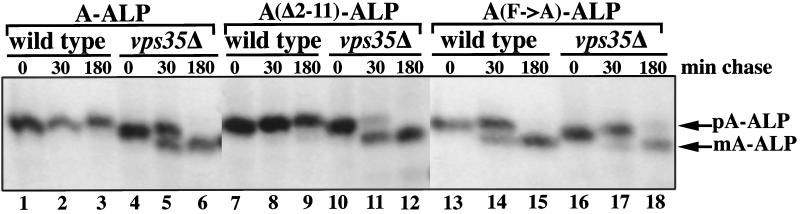
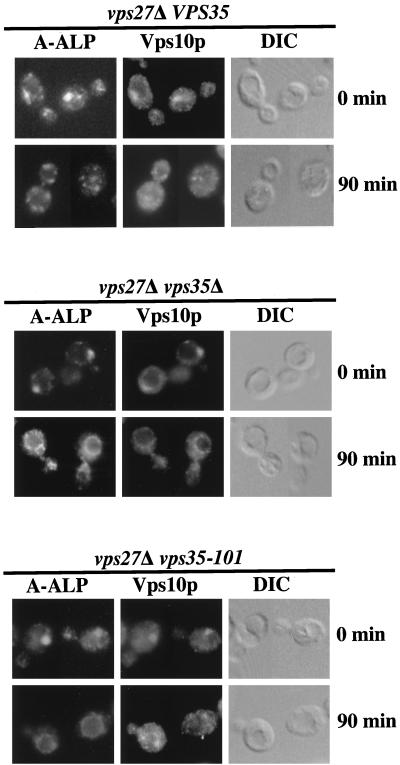
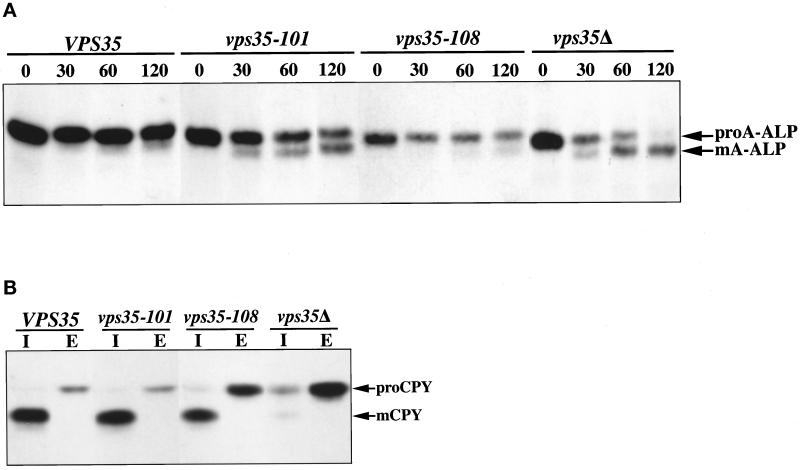
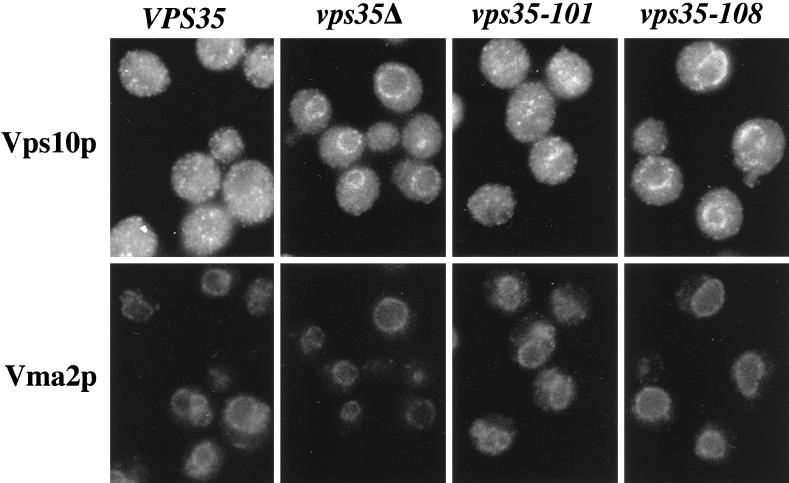
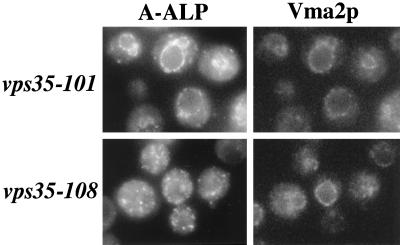
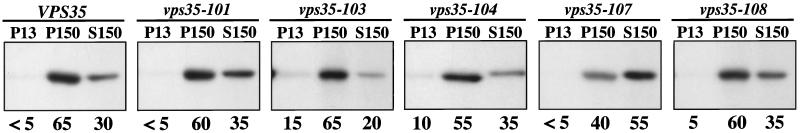
Resident membrane proteins of the trans-Golgi network (TGN) of Saccharomyces cerevisiae are selectively retrieved from a prevacuolar/late endosomal compartment. Proper cycling of the carboxypeptidase Y receptor Vps10p between the TGN and prevacuolar compartment depends on Vps35p, a hydrophilic peripheral membrane protein. In this study we use a temperature-sensitive vps35 allele to show that loss of Vps35p function rapidly leads to mislocalization of A-ALP, a model TGN membrane protein, to the vacuole. Vps35p is required for the prevacuolar compartment-to-TGN transport of both A-ALP and Vps10p. This was demonstrated by phenotypic analysis of vps35 mutant strains expressing A-ALP mutants lacking either the retrieval or static retention signals and by an assay for prevacuolar compartment-to-TGN transport. A novel vps35 allele was identified that was defective for retrieval of A-ALP but functional for retrieval of Vps10p. Moreover, several other vps35 alleles were identified with the opposite characteristics: they were defective for Vps10p retrieval but near normal for A-ALP localization. These data suggest a model in which distinct structural features within Vps35p are required for associating with the cytosolic domains of each cargo protein during the retrieval process.
Figures
References
-
- Abeliovich H, Grote E, Novick P, Ferronovick S. Tlg2p, a yeast syntaxin homolog that resides on the Golgi and endocytic structures. J Biol Chem. 1998;273:11719–11727. - PubMed
-
- Bednarek SY, Ravazzola M, Hosobuchi M, Amherdt M, Perrelet A, Schekman R, Orci L. COPI- and COPII-coated vesicles bud directly from the endoplasmic reticulum in yeast. Cell. 1995;83:1183–1196. - PubMed
-
- Boeke JD, LaCroute F, Fink GR. A positive selection for mutants lacking orotidine-5′-phosphate decarboxylase activity in yeast: 5-fluoroorotic acid resistance. Mol Gen Genet. 1984;197:345–346. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
