

Sla1p is a functionally modular component of the yeast cortical actin cytoskeleton required for correct localization of both Rho1p-GTPase and Sla2p, a protein with talin homology
- PMID: 10198057
- PMCID: PMC25233
- DOI: 10.1091/mbc.10.4.1061
Sla1p is a functionally modular component of the yeast cortical actin cytoskeleton required for correct localization of both Rho1p-GTPase and Sla2p, a protein with talin homology
Abstract
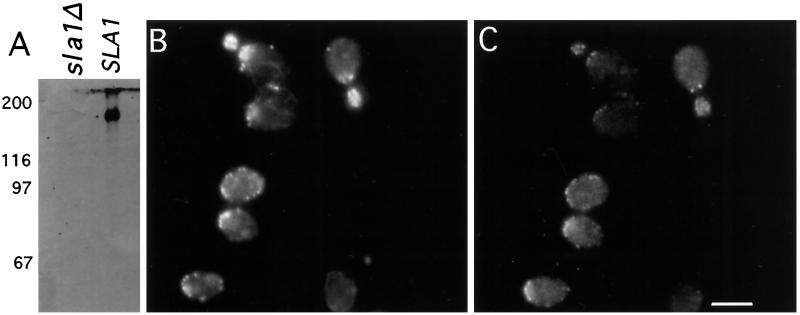
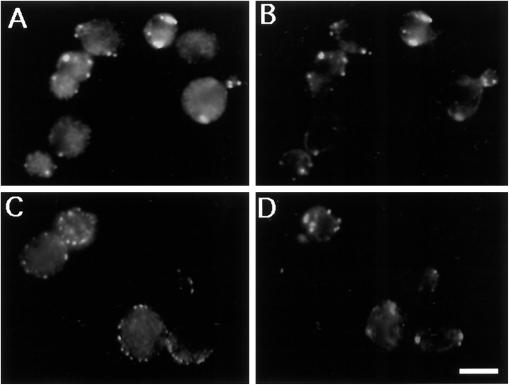
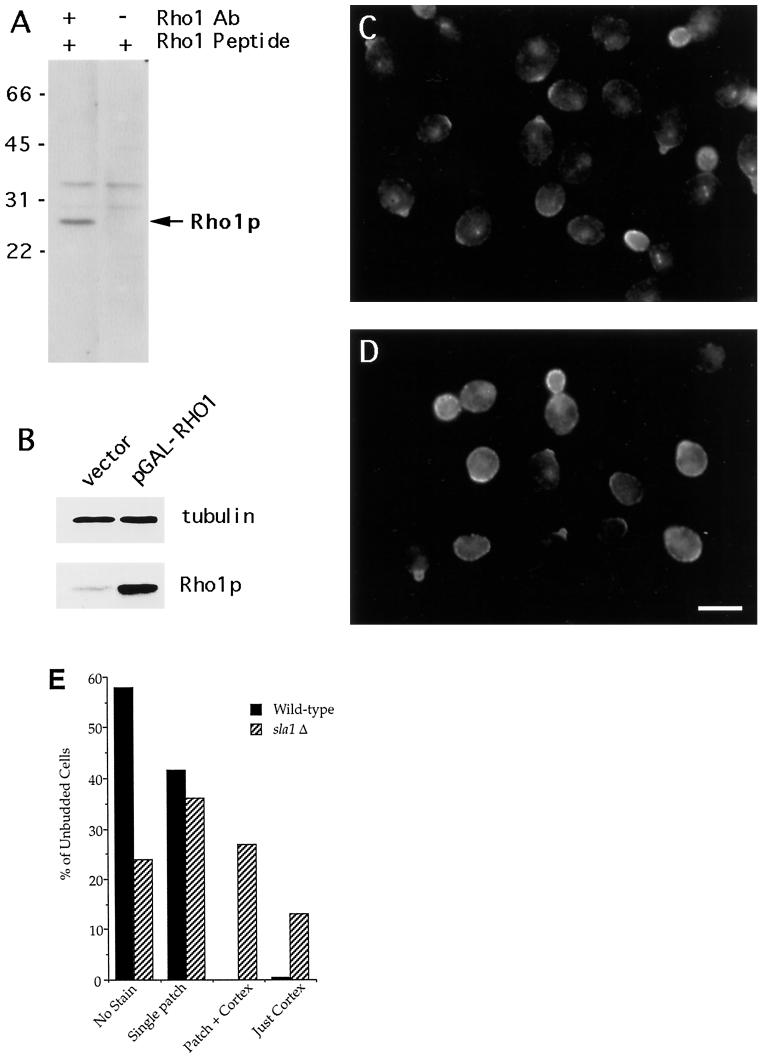
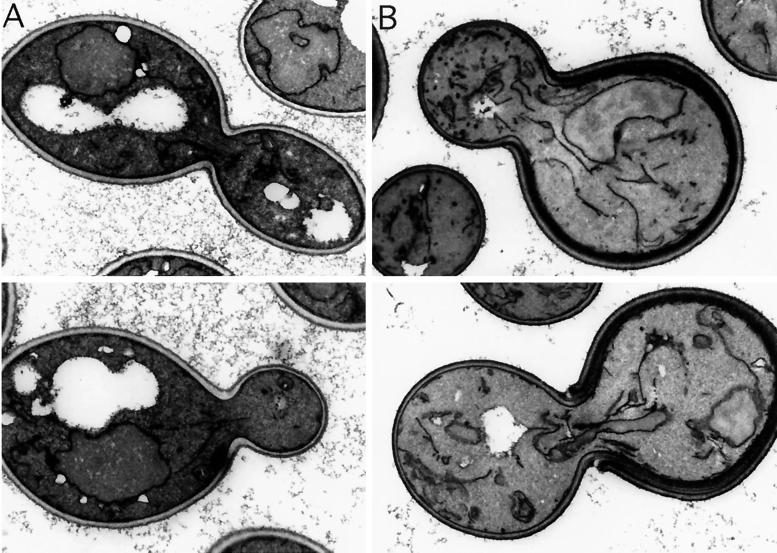
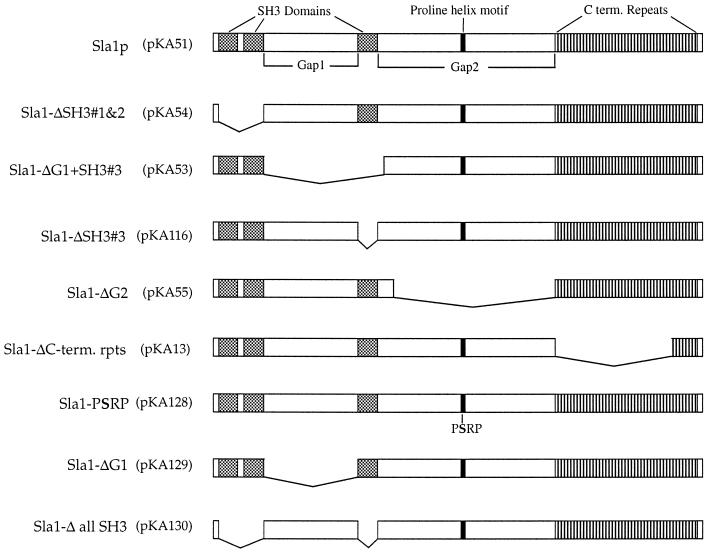
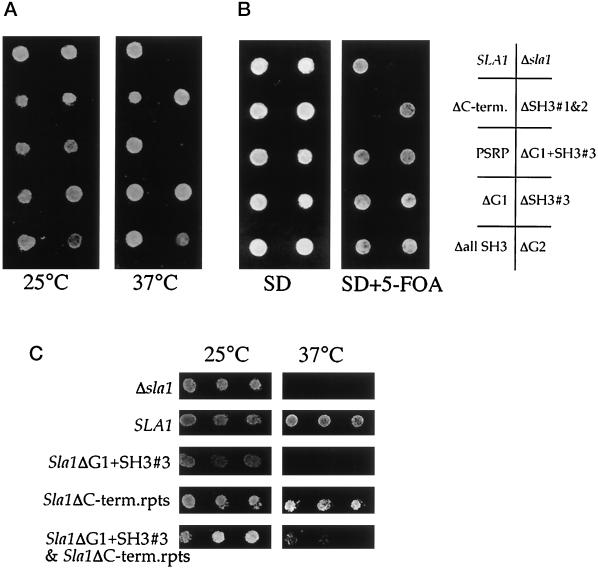
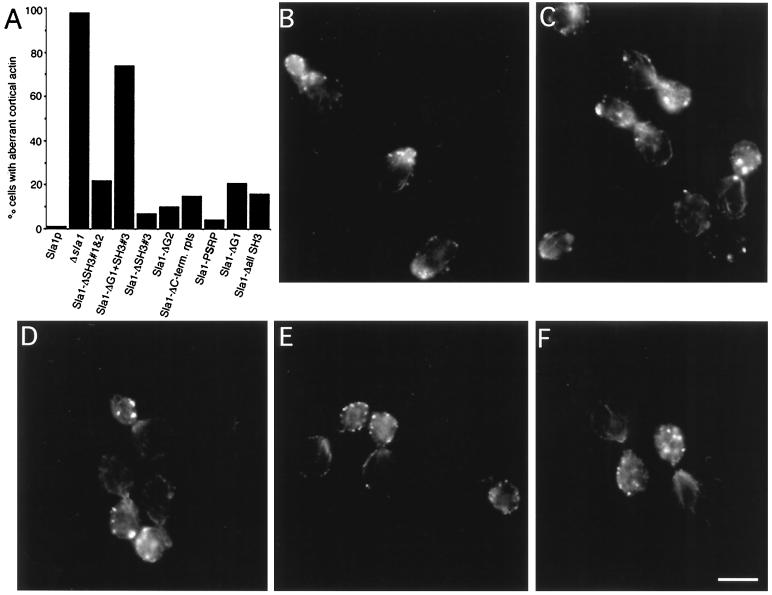
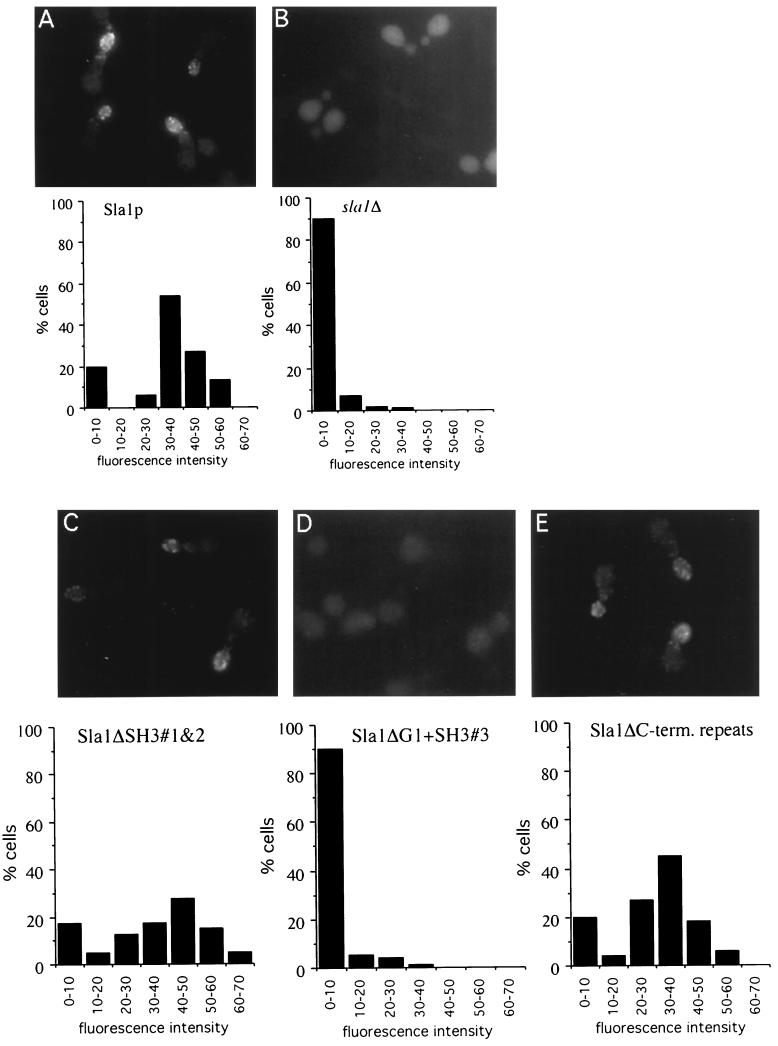
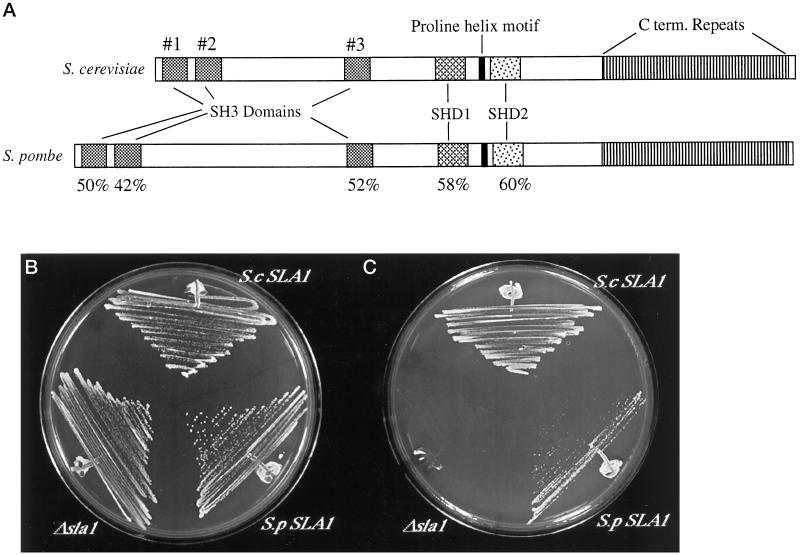
SLA1 was identified previously in budding yeast in a genetic screen for mutations that caused a requirement for the actin-binding protein Abp1p and was shown to be required for normal cortical actin patch structure and organization. Here, we show that Sla1p, like Abp1p, localizes to cortical actin patches. Furthermore, Sla1p is required for the correct localization of Sla2p, an actin-binding protein with homology to talin implicated in endocytosis, and the Rho1p-GTPase, which is associated with the cell wall biosynthesis enzyme beta-1,3-glucan synthase. Mislocalization of Rho1p in sla1 null cells is consistent with our observation that these cells possess aberrantly thick cell walls. Expression of mutant forms of Sla1p in which specific domains were deleted showed that the phenotypes associated with the full deletion are functionally separable. In particular, a region of Sla1p encompassing the third SH3 domain is important for growth at high temperatures, for the organization of cortical actin patches, and for nucleated actin assembly in a permeabilized yeast cell assay. The apparent redundancy between Sla1p and Abp1p resides in the C-terminal repeat region of Sla1p. A homologue of SLA1 was identified in Schizosaccharomyces pombe. Despite relatively low overall sequence homology, this gene was able to rescue the temperature sensitivity associated with a deletion of SLA1 in Saccharomyces cerevisiae.
Figures
References
-
- Ayscough KR. In vivo functions of actin-binding proteins. Curr Opin Cell Biol. 1998;10:102–111. - PubMed
-
- Ayscough KR, Drubin DG. Immunofluorescence microscopy of yeast cells. In: Celis J, editor. Cell Biology: A Laboratory Handbook. Vol. 2. San Diego: Academic Press; 1997. pp. 477–485.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
