

Modality-specific hyper-responsivity of regenerated cat cutaneous nociceptors
- PMID: 10200435
- PMCID: PMC2269289
- DOI: 10.1111/j.1469-7793.1999.0897u.x
Modality-specific hyper-responsivity of regenerated cat cutaneous nociceptors
Abstract
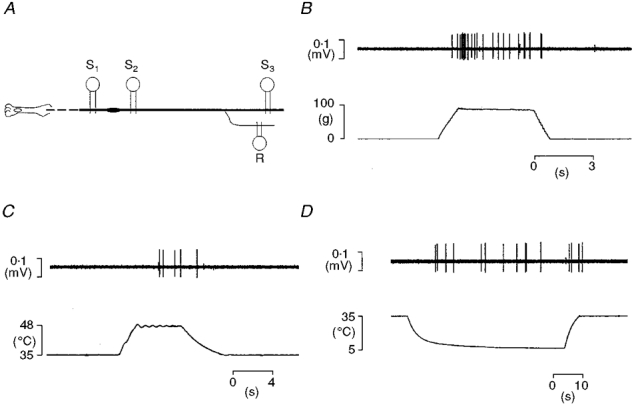
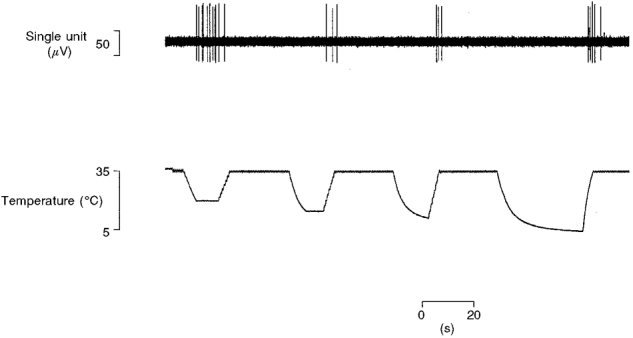
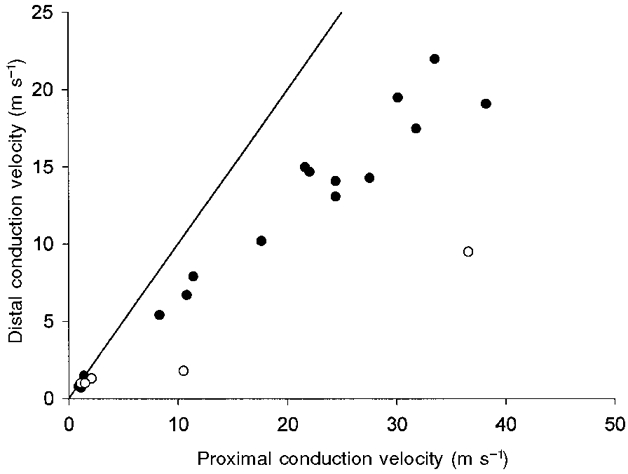
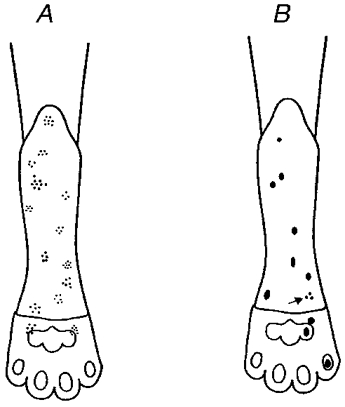
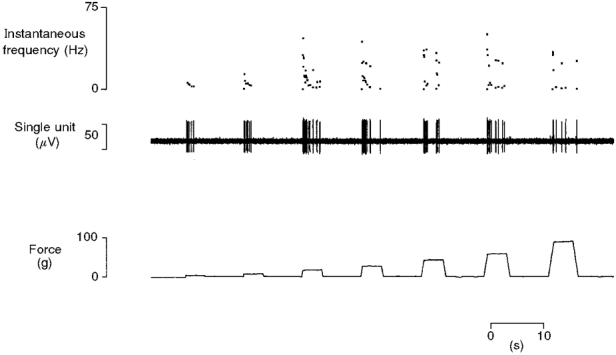
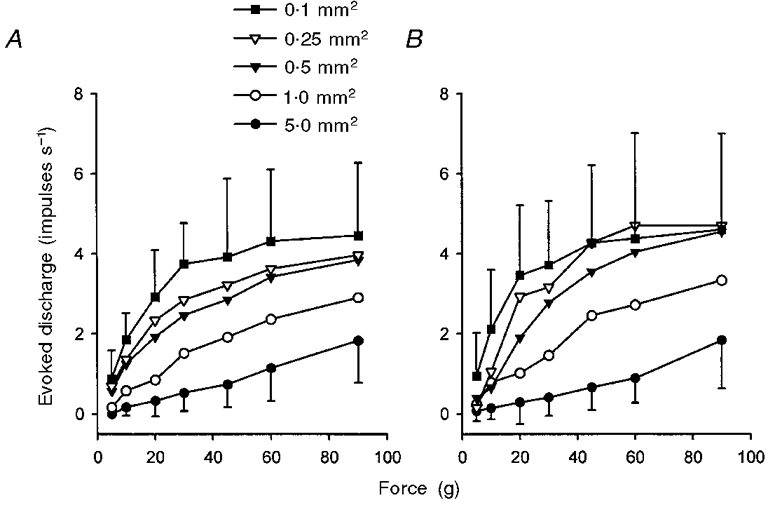
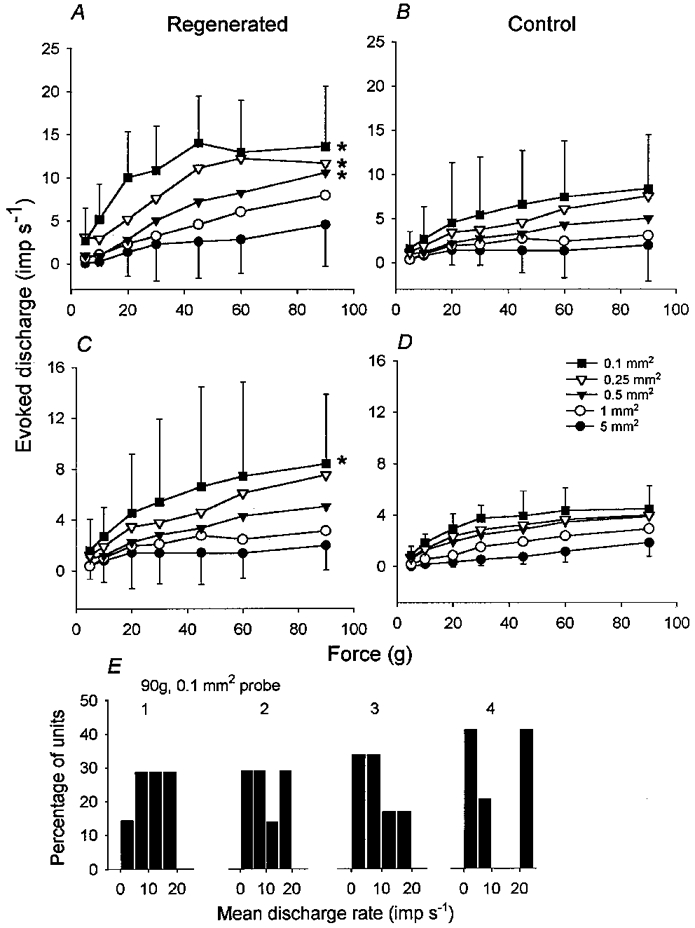
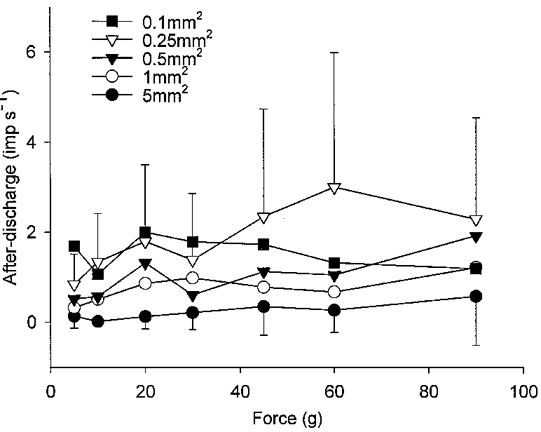
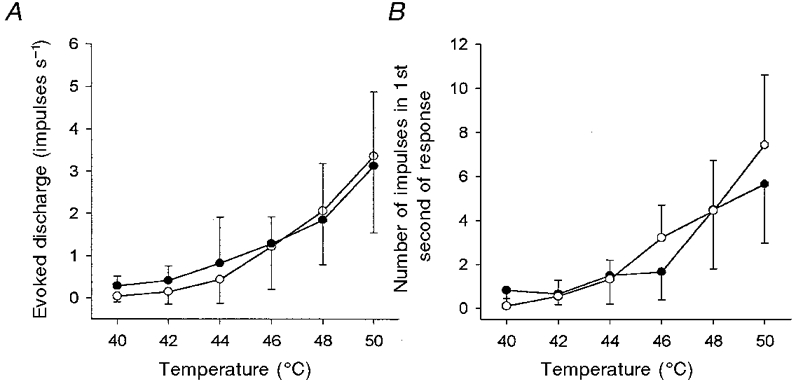
1. Experiments were performed on anaesthetized cats to investigate the receptive properties of regenerated cutaneous tibial nerve nociceptors, and to obtain evidence for coupling between them and other afferent fibres as being possible peripheral mechanisms involved in neuropathic pain. These properties were studied 6-7 months after nerve section and repair. 2. Recordings were made from 25 regenerated nociceptors; 14 were A fibres and the remainder were C fibres. Their receptive field sizes and conduction velocities were similar to controls. There was no significant difference between their mechanical thresholds and those of a control population of nociceptors. 3. Regenerated nociceptors were significantly more responsive to suprathreshold mechanical stimuli than were uninjured control fibres. This increase in mechanical sensitivity occurred in both A and C fibres, although A fibres showed a greater increase in mechano-sensitivity than C fibres. Over half of the regenerated nociceptors (13/25) showed after-discharge to mechanical stimuli which was never seen in controls; the mean firing rate during this period of after-discharge was significantly related to both stimulus intensity and stimulus area. 4. There was no significant difference between the heat encoding properties of regenerated nociceptors and control nociceptors. Cold sensitivity was similarly unchanged. Thus, abnormal peripheral sprouting was unlikely to account for the increased mechanical sensitivity of the regenerated fibres. None of the regenerated nociceptors were found to be coupled to other fibres. 5. These results suggest that the clinical observation of mechanical hyperalgesia in patients after nerve injury may have a peripheral basis. Based on this model, other signs of neuropathic pain (i.e. tactile or thermal allodynia) are more likely to be due to altered central processing.
Figures

References
-
- Ahlgren SC, White DM, Levine JD. Increased responsiveness of sensory neurones in the saphenous nerve of the streptozotocin-diabetic rat. Journal of Neurophysiology. 1992;68:2077–2085. - PubMed
-
- Beck PW, Handwerker HO, Zimmerman M. Nervous outflow from the cat's foot during noxious radiant heat stimulation. Brain Research. 1974;67:373–386. - PubMed
-
- Bennett GJ. Neuropathic Pain. In: Wall PD, Melzack R, editors. Textbook of Pain. New York: Churchill Livingstone; 1994. pp. 201–224.
-
- Bessou P, Perl ER. Responses of cutaneous sensory units with unmyelinated fibres to noxious stimuli. Journal of Neurophysiology. 1969;32:1025–1043. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Miscellaneous